II.7.1 Trait heritability can be estimated by establishing the level of correlation between the characteristics of parents and their offspring or by observing the phenotype response to selection pressure.
There are two basic procedures for determining the degree of heritability of a trait (h2). Francis Galton, a cousin of Charles Darwin, used the first of these in the 19th century. This consists in the determination of the correlation between the mean value of a particular trait in both parents and the value of this trait in their progeny (Fig. II.16).
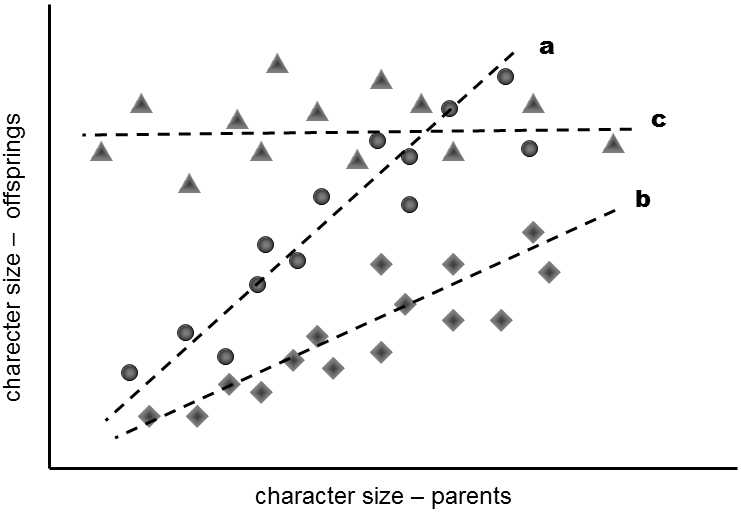
Fig. II.16 Determination of the heritability of quantitative traits by the regression method. If we plot the values of a trait for a certain individual against the average values for the same trait in his parents, we can roughly draw a straight line through the points. The greater the Pearson coefficient for the correlation of the given straight line, the greater is the heritability of the given trait.
As the genome of any descendant contains only half the genes (or, to be more exact, alleles) of all the genes (alleles) contained in the genomes of the parents, it is necessary to multiply the obtained correlation coefficient by two to obtain the calculated heritability coefficient h2. In order to reduce the effect of the nonadditive component of the heritability on the observed correlation between the offspring and the parents, the given correlation is studied in practice, not on the basis of measuring the correlation between the characteristics of the parents and their progeny, but rather on the basis of correlation between the characteristics of the siblings sharing only one parent. Genes derived from the common parent always occur in each offspring together with other genes so that any interactions amongst genes derived from the mother and from the father cannot participate in the correlation (but gene interactions amongst genes of the same parent can, see below). In this arrangement, the compared individuals, i.e. half-siblings, share only ¼ of their genes, and thus the measured correlation coefficient must be multiplied by four to obtain calculated heritability coefficient.Galton studied the heritability of body height and found two important facts. To begin with, he demonstrated that heritability in a given trait does actually exist, as tall parents actually tend to have tall children and short parents tend to have short children. Secondly, he formulated the law of regression to the mean (Bateson 2001; Lewontin 1974). The further the mean height of the parents from the population mean, the greater was the probability that the height of their children would return back towards the population mean, rather than deviating even further from this mean than the deviation of the mean height of their parents. This return towards the mean can be explained by the existence of non-additive components of genetically determined variability. In a stabilized population, the population mean of the value of a quantitative characteristic should correspond to the optimal value of this trait from the standpoint of the biological fitness of its carriers. As individuals with larger or smaller values of the given trait are constantly removed from the population by normalized selection, a frequency of the individual alleles of the genes affecting the given trait is established in the gene pool of the population that leads to the optimal value of the given trait in the largest possible number of random combinations. If the heights of the mother and of the father deviate substantially from the mean, they most probably have some rare combination of the relevant alleles. This combination will disappear in their progeny either immediately or in the subsequent generations. If only genes with additive effect were relevant for the given trait, the progeny should not return to the population mean.
Galton could, of course, not correctly interpret the results of his study under the conditions at that time. He explained the existence of regression to the population mean as an indication that the characteristics of an individual are determined 50% by predispositions obtained from their parents, 25% by predispositions obtained from their grandparents, 12.5% from their great-grandparents, etc. If predispositions are interpreted as genes in the sense of cistrons, then this would be a quite erroneous explanation, as an individual obtains all his genes from his parents. However, if we realize that a predisposition can also be the effect of certain interactions of a combination of several alleles at various loci, a combination that an individual inherits from his predecessors but that can fall apart or, to the contrary, be formed with a certain probability in each generation, then this, at first glance erroneous explanation, can be basically correct. The probability that an individual inherited a certain combination of alleles from one of his great grandparents is less than the probability that he inherited it from his grandparents and this is again less than the probability that he would inherit it from one of his parents.
The second approach to determining the heritability of a trait consists in its calculation from the magnitude of the response of the population to directional selection of a certain known intensity, i.e. the magnitude of the shift in the population mean of the value of the given trait from one generation to the next (Fig. II.17). The intensity of the selection can be expressed as the selection differential, i.e.
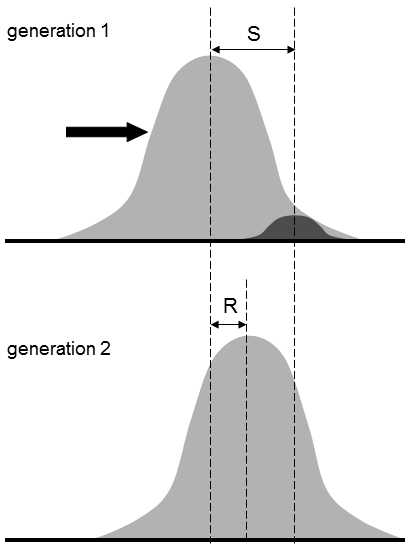
Fig. II.17 Determination of the heritability of quantitative traits using a selection experiment. If we subject a population to directional selection, through which the average value of a certain trait is shifted relative to the value prior to commencing the selection by value S, the average value of the trait will most probably return in the subsequent generation to a value somewhere between the value prior to commencing the selection and that after its completion. This shift in the average value of the trait compared to the original value is termed the evolutionary response and is denoted by symbol R. Ratio R/S is numerically equal to the additive heritability of the particular trait.
the difference in the value of the population mean for the given trait prior to and after the selection. The degree of evolutionary response of the population to selection can be derived from the shift in the value of the population mean between two generations. If the evolutionary response is divided by the selection differential, then the coefficient of heritability of the given trait (h2) is obtained.
It must be stressed, however, that all normally employed models of heritability are greatly simplified and thus they do not describe this phenomenon to its full extent. This leads, amongst other things, to the unpleasant fact that estimates of the heritability of the individual traits by the methods of measuring the correlation between the characteristics of parents and their offspring and monitoring the phenotype response to selection pressure can frequently differ very substantially (Lynch & Walsh 1998).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.