IV.9.2 The theory of frozen plasticity assumes that species of sexually reproducing organisms exhibit evolutionary plasticity only immediately after emergence, before genetic polymorphism builds up in their genetic pool.
The main drawback of the selfish gene theory is that it does not take into account the existence of epistatic interactions between individual genes and the consequent dependence of the phenotype manifestations of the individual alleles on their own frequency and on the frequency of alleles in other loci.The model of inter-allele selection tacitly (and erroneously) assumes that the effects of the individual genes on the phenotype of the organism and, indirectly, on the biological fitness, are simply additive.In actual fact, this is frequently not true (Chippindale & Rice 2001). Two alleles, each of which can be advantageous on its own for its bearer, can frequently substantially reduce the biological fitness of their bearer if they are in the same genome.On the other hand, two independently harmful alleles can, together, increase the fitness of their bearer.This means that the fact that the actual allele (in contrast to the overall genotype) is inherited from one generation to the next in unaltered form still does not ensure that its phenotype manifestations and their effect on the biological fitness of the individual will also be inherited (Fig. IV.13) and that they can spread through the Darwinistic mechanism of natural selection.
For example, in most individuals in the human population, the effect of loss mutations on the gene encoding the α-chain of haemoglobulin is highly negative, as inadequate production of this chain causes α-thalassemia.However, in bearers of a similar mutation in the gene for the β-chain of haemoglobulin, the same mutation is manifested in an increase in their biological fitness, as it prevents the occurrence of relative excessive production of the α-chain and thus also the occurrence of pernicious β-thalassemia (Wainscoat et al. 1983; Kanavakis et al. 1982).Thus we cannot assign any specific selection coefficient value to the individual alleles.In addition, even if we could do this,
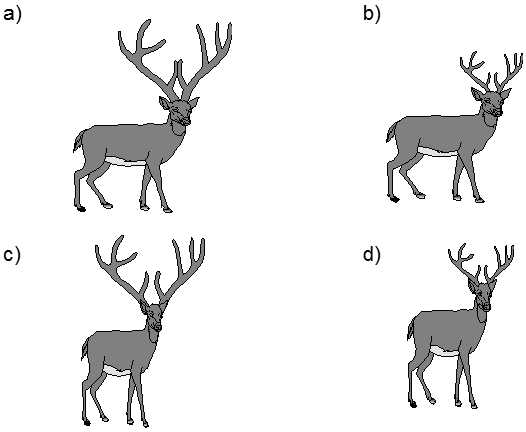
Fig. IV.13. Context determination of the effect of individual traits on the biological fitness of the bearers. In this case, enormous antlers are advantageous for the normal form of elk, as they ensure a greater chance of success in the battle for females. However, larger antlers tend to be a handicap for elk with less robust stature, they will lose in combat anyway and, in addition, the antlers make them clumsy
the evolutionary fate of the alleles will not be determined by the value of such a coefficient, but rather by whether the allele determines an evolutionarily stable strategy (see IV.5.1.1).Thus the solution to the problem of inadequate heritability of biological fitness suggested by Dawkins in his selfish gene theory (the model of interallelic selection) is inadequate.
As a single mutation occurs in the context of other genes in each generation as a consequence of mixing of the genomes in sexual reproduction and its effect on the properties of the bearer thus changes, a major proportion of mutations cannot become fixed in the gene pool of the species.Thus, sexually reproducing species gradually begin to exhibit an increasing degree of genetic polymorphism.Increased polymorphism again increases the probability that the new mutation will find itself in the context of a different gene pool in each generation.This viscous circle of positive feedback, together with a common phenomenon of frequency dependent selection (see chapters IV.5, IV.5.1 and IV.5.1.1), finally leads to the formation of genetic homeostasis, a phenomenon that was recognized long ago in their experiments by classical geneticists (Lerner 1958).Initially, a species changes readily under the effect of any selection pressure; however, as the frequency of the individual alleles gradually shifts away from equilibrium, the selection coefficients of the individual alleles present also gradually changes until the selected population ceases to respond to the relevant selection pressure (Fig. IV.14).This phenomenon is sometimes interpreted in that, during the selection experiment, the genetic variability in the population is exhausted and further evolution of the trait begins to be limited by waiting for a new mutation.This explanation is, however, apparently erroneous.If, at the moment when the evolution of the given trait stops, we begin to exert pressure the opposite direction, for example, instead of individuals with large dimensions of the given trait, we begin to select individuals with small dimensions of the trait, the population begins to respond quite willingly to the new pressure and the relative trait will become smaller.However, this means that, at the time when the population does not react to the original selection pressure, it still contained the genetically-dependent variability.The most probable explanation for genetic homeostasis consists in pleiotropic negative effects on the genes responsible for the given trait.Some alleles that became fixed through selection pressure of the experimenter, or at least increased their frequency, simultaneously negatively affect the biological fitness of their bearer.This means that, from a certain instance, the bearers of a particular combination of alleles continue to be at an advantage through the artificial selection of the experimenter, but are increasingly at a disadvantage in relation to natural selection.At the moment when the artificial selection and natural selection become equal, the development of the given trait stops.After termination of the selection pressure of the experimenter, a sufficiently large population will return to the original state through the action of natural selection, and the original frequency of the individual alleles and also the original phenotype (appearance and behaviour) of the individuals in the population are renewed in the population.In a small population, the return to the original state need not be complete, as some less frequent alleles disappear from the population through drift (see chapter V).
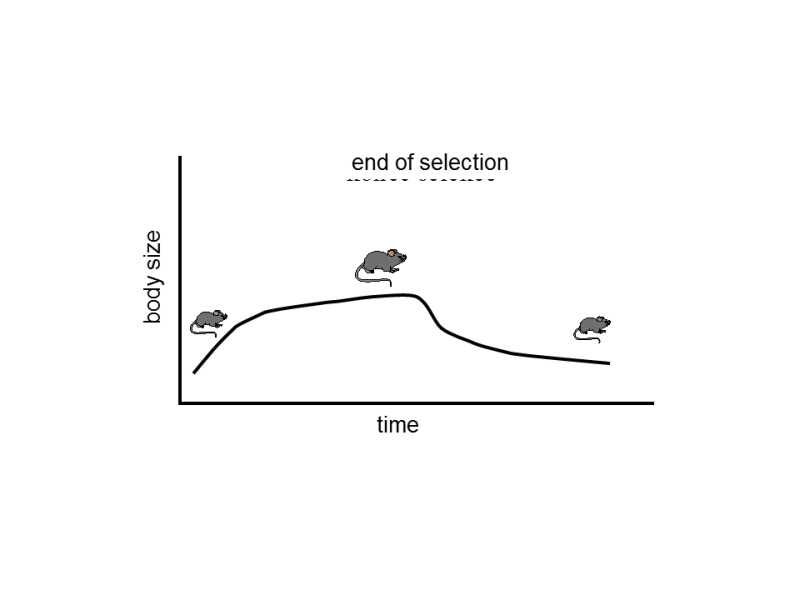
Fig. IV.14. Genetic homeostasis. If a population is artificially subjected to directional selection for a particular trait, in this case for greater body weight, after a certain time there will be a slowing or even complete stopping of the evolutionary response to the selection pressure. After the end of the experiment, the value of the trait can spontaneously return to the original value. The return to the original value is caused by the fact that natural selection returns, to the original value, the frequency of alleles that, while they were responsible for the greater body weight of mice, and were thus preferred in artificial selection on the part of the experimenter, simultaneously reduced the fertility or viability of their bearers. The progress of the experiment indicates, amongst other things, that, at the time when the population practically ceased to respond to the selection pressure, its gene pool continued to contain a large amount of genetic variability capable of affecting the relevant trait.
The theory of frozen plasticity (Flegr 1998),)assumes that, as a consequence of the occurrence of genetic homeostasis, clearly punctuated evolution (see XXVI.5) is characteristic for sexually reproducing species.Throughout most of their existence, species more or less do not change or change only temporarily, in spite of frequently dramatic changes in their environment.Irreversible changes in the properties of a species, i.e. anagenesis, occur only immediately following speciation, when the size of originally small population of the newly formed species has already grown (therefore the destiny of an individual is already directed by selection, not a chance, see chapter V.4) but the population still bears only a small fraction of the polymorphism of the parent species.The drastic reduction in the genetic polymorphism in the new species means that new mutations are present in the company of the same genes even in sexual reproduction.Until sufficient genetic polymorphism accumulates in the gene pool of the population, the species is evolutionarily plastic and can respond adaptively to selection pressures of the environment similarly to asexual species.Following accumulation of polymorphism, the species evolutionarily “freezes” (becomes evolutionary frozen on macroevolutionary time-scale and evolutionary elastic on microevolutionary time-scale) and, for the rest of its existence, only passively waits until a change in its environment causes its extinction.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.