VI.1.1.1 Mutation bias and reparation drive can be responsible for the emergence of isochores in the genomes of warm-blooded organisms.
The genomes of warm-blooded vertebrates, i.e. mammals and birds, are characterized by the presence of isochores. These are a DNA segments that are 0.2 to 1.3 million nucleotides long and that differ in their contents of G-C pairs. Segments rich in G-C pairs are called H-isochores (H for Heavy – the heavy fraction of DNA in isopycnic centrifugation). In mammals and birds, the H-isochores encompass 35-50% of the genome. Segments poor in G-C pairs are called L-isochores (L for light – the light DNA fraction). The function of isochores and why they are encountered in warm-blooded vertebrates are not yet known with any certainty. One group of hypotheses assumes that they could be formed as a product of natural selection, possibly in connection with the necessity of adapting the mean melting point of DNA to the elevated body temperature (Bernardi 2000). Codons with a high content of G-C pairs also code primarily hydrophobic aminoacids that exhibit higher thermodynamic stability (Alvarez-Valin, Lamolle, & Bernardi 2002). An alternative explanation lies in the joint action of mutation bias and reparation drive. In 2000, Fryxell a Zuckerkandl (Fryxell & Zuckerkandl 2000)suggested a very probable model for the formation of isochores. According to this model (see Fig. VI.2) isochores are formed as a
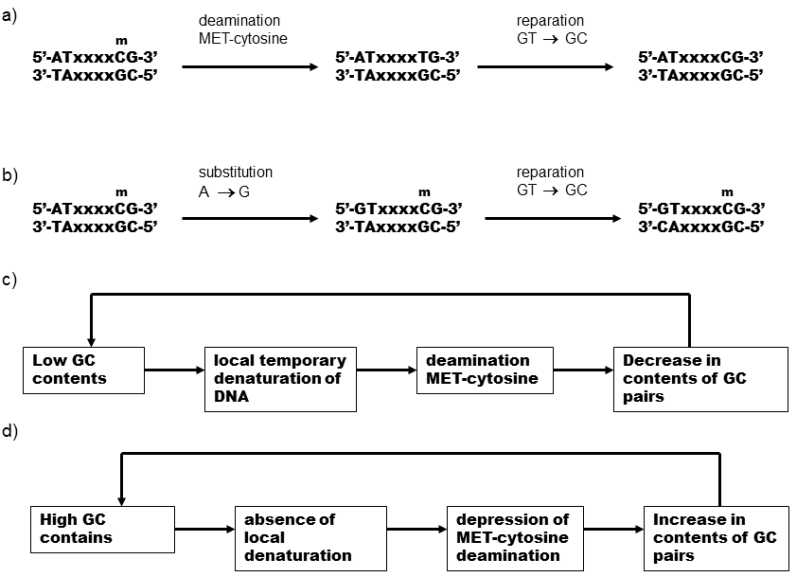
Fig. VI.2. Model of the formation of the isochore structure of the genomes of warm-blooded organisms. Three types of mutations affecting the content of GC-pairs may occur in the DNA (amongst other things): 1) deamination of methyl cytosine leads to the formation of thymines, reducing the content of GC-pairs (a), 2) substitution of T for C and 3) substitution of C for T (b). Deamination of the bases occurs very readily in warm-blooded organisms. Consequently, evolution has developed a strong tendency of the reparation apparatus to preferentially replace T in nonpairing TG couples by cytosine, which is capable of neutralizing the effect of mutation bias. The strength of the reparation drive is adjusted by evolution to a value that exactly compensates the average intensity of mutation bias, acting in the opposite direction. However, the individual DNA sections (future isochores) differ in their mutation bias intensity. If fewer GC-pairs and more AT-pairs (which are coupled only by two hydrogen bonds) are present at a certain DNA section, temporary local denaturing of the DNA occurs here more readily and more frequently. Deamination of the base and subsequent replacement of methyl cytosine by thymine, occurs far more readily in the denatured sections. Reparation drive is optimized from the standpoint of the average intensity of mutation bias, so that it is not capable of fully compensating its effect in sections with subnormal content of GC-pairs. Consequently, at this site, cytosine is gradually replaced by thymine, with a further reduction in the content of GC-pairs and thus ever more frequent deamination of the base occurs. The positive feedback cycle is thus closed, leading to a substantial reduction in the content of GC-pairs at this site (c). In contrast, in sections with above-average content of GC pairs, a similar positive feedback cycle leads to a further increase in the content of GC-pairs. At these sites, reparation drive acts more strongly than is required for compensation of mutation bias and thus erroneously replaces an increasing number of incorrectly placed adenines by guanines (d).
result of the action of mutation bias, reparation drive and because of the existence of positive feedback between the content of G-C pairs in the DNA strand and the probability of mutations leading to a further increase in the content of G-C pairs. The commonest mutation in warm-blooded organisms is deamination of cytosine and 5-methyl cytosine that leads to replacement of cytosine by thymine and subsequently, in the opposite site, replacement of guanine by adenine. These mutations occur with more than 140-fold probability in denatured, i.e. single-strand DNA segments, i.e. particularly in segments where there is a low content of G-C pairs, i.e. pairs of bases connected by three hydrogen bonds and where temporary local denaturing of DNA, called DNA breathing, occurs more readily. Thus, if random fluctuations in a certain DNA segment leads to a reduction in the content of G-C pairs, there is increased probability of deamination of cytosine and 5-methyl cytosine and thus increased probability of the occurrence of mutations leading to a further decrease in the number of G-C pairs in the given segment. In segments where, on the other hand, random fluctuations lead to an increase in the content of G-C pairs, the situation is analogous and positive feedback again augments the given fluctuation. In this case, the mechanism of strengthening fluctuations is somewhat more complicated, with participation by reparation drive. Here the nonpairing GT pair is preferentially repaired to GC (approx. 24x more frequently) and not toAT in warm-blooded vertebrates (Brown & Jiricny 1988). In the nondenatured segments, e.g. in segments rich in G-C pairs, the rate of deamination of cytosine and methyl cytosine is low and couples of nonpairing GT bases are formed here with approximately the same probability as replacement of nucleotide A by G as replacement ofC byT. Preferential repair of T to C at this site thus leads to a gradual increase in the content of G-C pairs
The intensity of deamination is highly dependent on the temperature; an increase in temperature by 10 oC leads to an almost six-fold increase in the reaction rate. This means that, in warm-blooded organisms, processes accompanying deamination of DNA are of extraordinary importance and can thus decisively affect the formation of isochore structures of the genome.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.