VI.3.1 An allele is very often able to influence whether it will end up in the egg nucleus or in the polar body during female gamete development.
Meiotic drive occurs very frequently in egg formation. During this process, only one haploid set of chromosomes ends up in the nucleus of the female gamete, while the remaining three sets are eliminated into the polar bodies. In heterozygote females, it very frequently occurs that the probability with which a certain allele will end up in the nucleus of the oocyte or in the polar body, i.e. the probability with which the allele will be transferred to the next generation, differ considerably (Fig. VI.10). (Crow 1979). In some cases a certain allele of a specific gene is proliferated in this way in the
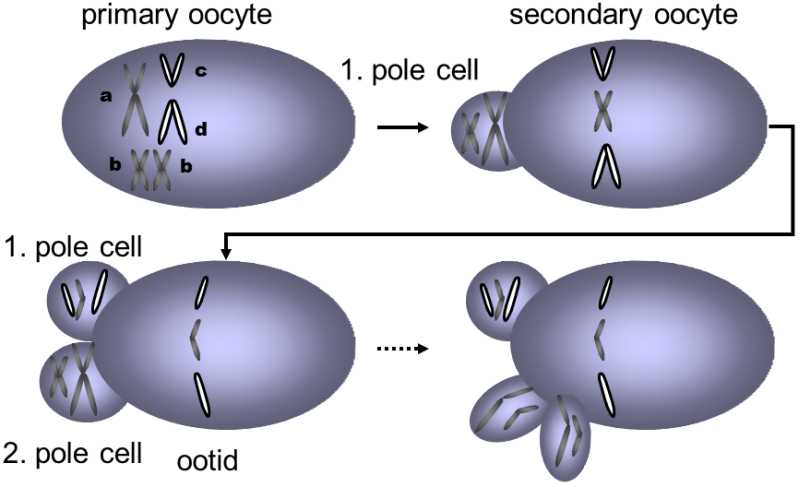
Fig. VI.10. Meoitic drive in the oocyte. In reduction division in the forming oocyte, only one of the four chromatids of the two homologous chromosomes of the original tetraploid cell ends up in the nucleus of the oocyte. The other three end up in the polar bodies and are not passed on to future generations. Under normal circumstances, chance should decide on the fate of the individual chromosomes and the allele of chromosome form should end up in the oocyte or in the polar body with the same probability. However, in some cases, a certain allele or a certain chromosome form almost always ends up in the nucleus of the oocyte and only in rare cases in the polar body. In this case, this allele or chromosome form can spread faster in the population through meiotic drive. In the figure, telocentric chromosomes c and d, which were apparently originally formed by splitting of chromosome a in the area of the centromere, are given an advantage by meiotic drive compared to metacentric chromosome a.
population while, in a different case, this can be a certain chromosome mutation (Ruvinsky 1995). If, for example, a laboratory mouse is crossed with a wild house mouse, whose karyotype contains a metacentric chromosome formed through Robertson translocation, i.e. fusion of two acrocentric chromosomes, it has been observed in five cases out of ten that the metacentric chromosome occurs in less than 50% of the progeny of heterozygote females. In some cases, the ratio of the two types of oocyte was as much as 3:1. The authors assumed that the metacentric chromosome would most probably end up in the primary polar body. In another study, monitoring the behaviour of a metacentric chromosome in a population of wild mice, it was observed, on the other hand, that the metacentric chromosome had the greatest probability of ending up in the nucleus of the oocyte (King 1993).
Similar phenomena were also observed for other organisms. For example, it has been observed in sorrel (Rumex acetosa) that only four of nine randomly selected chromosome mutations of the reciprocal translocation type or mutations externally manifested as a shift in centromers exhibited normal Mendelian heredity. In the remaining five, meoitic drive appeared to some degree, in the female or the male plants.
DNA sequencing in the area of the centromere (the site of attachment of microtubules of meiotic spindle and therefore probably an important battlefield for meiotic distorters) in closely related species indicated that these areas are subject to very rapid evolution. It is highly probable that this is the result of a battle between genetic elements proliferating through meoitic drive. According to some authors, a significant part of the DNA in the area around the centromere is formed of these active or inactive elements – meiotic distorters. The predominance of nonsynonymous mutations in the histones that are bonded to the DNA in the area of the centromere, is interpreted in a similar way (a battle between meiotic distorters).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.