VIII.4.1 Stable persistence of recessive harmful alleles with very high frequency in a population can be the consequence of relatively low mutation pressure
As we have already demonstrated in the section on genetic models of natural selection (see IV.6.1), selection against a rare recessive allele is rather ineffective.For example, if the frequency of the recessive allele is 5%, then only 0.25% of the individuals in the population will form recessive homozygotes and only these individuals will be exposed to selection pressure.Recessive alleles frequently code a biologically inactive protein.These alleles can be formed very readily and thus very frequently by mutation from dominant functional alleles.Repeated formation of the same allele as a consequence of mutation is termed mutation pressure.The probability of reverse mutation is much less than the probability of the original mutation and thus, as a consequence of mutation pressure, the number of copies of the mutated alleles increases in the population.In case of mutations reducing the fitness of an organism, selection acts in the opposite direction against mutation pressure and removes the mutated alleles from the population.However, if the frequency of the recessive alleles is low, the effectiveness of selection is also very low, so that these alleles can persist permanently in the gene pool of the population with unexpectedly high frequency (Fig. VIII.2a).The equilibrium frequency of recessive alleles in the population depends on the strength of
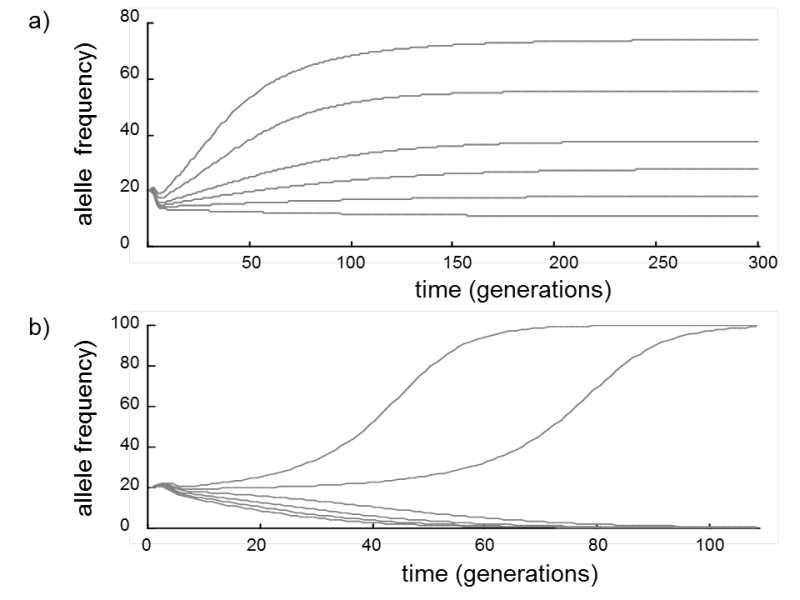
Fig. VIII.2. The effect of mutation recessivity on its survival in the population. If the harmful recessive allele a is formed from the dominant allele A repeatedly in the population through mutations, and these alleles are continuously removed by selection acting against their homozygote bearers, there is a wide range of values of the mutation rate and selection coefficient at which the mutated alleles survive in the population with relatively high frequency (a). However, if the given mutated alleles are dominant or semi-dominant, there is no combination of values of the mutation and selection pressure at which a stable equilibrium frequency of the mutated alleles would be maintained in the population. At high mutation pressure values, these alleles a would become fixed; at low values, their content in the population would be maintained at a value corresponding approximately to the number of new mutations in each generation (b). Model employed: the density of the population growth is
![]()
![]()
![]() , where NAA, NAa, Naa are the numbers of individuals of the particular genotype, rAA, rAa, raa are the growth rates of individuals of the particular genotype, (1.2, 1.2 and 1.1 for recessive mutations and 1,2, 1,1 and 1,1 for dominant), the probability of mutation conversion from allele A to allele a for a recessive allele (upper graph) is 0.01, 0.015, 0.02, 0.025, 0.030 and 0.035 and, for a dominant allele (lower graph), equals 0.05, 0.055, 0.06, 0.065, 0.07 and 0.075. The numbers of individuals of genotypes AA, Aa and aa at time t0 are equal to 32, 16 and 2.
, where NAA, NAa, Naa are the numbers of individuals of the particular genotype, rAA, rAa, raa are the growth rates of individuals of the particular genotype, (1.2, 1.2 and 1.1 for recessive mutations and 1,2, 1,1 and 1,1 for dominant), the probability of mutation conversion from allele A to allele a for a recessive allele (upper graph) is 0.01, 0.015, 0.02, 0.025, 0.030 and 0.035 and, for a dominant allele (lower graph), equals 0.05, 0.055, 0.06, 0.065, 0.07 and 0.075. The numbers of individuals of genotypes AA, Aa and aa at time t0 are equal to 32, 16 and 2.
mutation pressure, i.e. on the frequency with which recessive alleles are formed from normal dominant alleles during a single generation and on the strength of the selection pressure acting against the homozygous carriers of the mutated alleles.Under conditions where no form of regulation of the size of the population is active in the population, i.e. in classical population-genetic models, the equilibrium frequency of the mutated allele (Q) can be calculated from the equation

where u is the strength of the mutation pressure and s is the selection coefficient acting against recessive homozygotes.An analogous equation is also valid for cases where selection is directed against dominant homozygotes.If selection is directed against heterozygotes, the equilibrium frequency of the mutated alleles in the population is much lower – specifically it is approximately just equal to the mutation rate (Li 2003).
While recessive mutations can survive in the population in a stable equilibrium state for a wide range of parameters u and s, dominant and partly dominant harmful alleles become fixed at (unrealistically) high mutation pressure values and almost disappear from the population at other values (Fig. VIII.2b).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.