XIII.3.1.3.3 Diploidy may have played a major role in the emergence of multicellularity
In all multicellular organisms, the diploidal phase forms a substantial part of the life cycle.Some biologists assume that the emergence of diploidy was an essential condition for the emergence of multicellularity (Lewis & Wolpert 1979; Gorshkov & Makar'eva 1999).The connection between diploidy and multicellularity is not particularly obvious; however, it could consist in enforcedly different strategy of the regulation of gene expression in diploid and haploid organisms (Lewis & Wolpert 1979).
In haploid organisms, most regulation occurs through the cis-mechanism, i.e. the regulation area, at which evaluation (integration) occurs of frequently opposing signals derived from the internal and external environment of the cell, is right next to the regulated gene.On the other hand, in diploid organisms, most of the regulation occurs through a trans mechanism, i.e. regulation is mediated by a product of a regulator gene, frequently coded in a distant region of the genome, for example on a different chromosome.The signals affecting switching the regulated gene on and off are evaluated directly on this product (usually a regulation protein), frequently even before it bonds to the relevant area of the regulated gene.This is apparently because both copies of the gene must be regulated at once in diploid organisms.It would seem that the same goal, simultaneous regulation of both copies of the gene, could also be achieved through a cis-mechanism.Such a regulation mechanism would emerge and further develop through the action of natural selection only with great difficulty in diploid organisms (Fig. XIII.5).
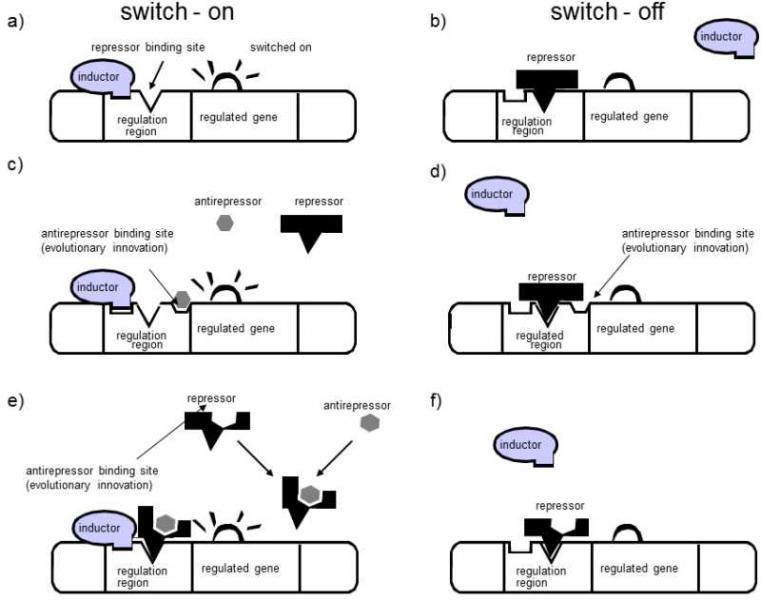
Fig. XIII.5 Preferential formation of a trans-mechanism of regulation of gene expression in diploids. Expression is originally regulated using an inducer, which switch on the transcription of the gene (a) and repressor that, through its presence, can block binding sites for the inducer and thus prevent commencement of transcription. The repressor has greater affinity for the DNA than the inducer so that, when the inducer and repressor are present simultaneously, gene expression will be switched off (b). In evolution, it can be advantageous if the particular gene is expressed in the presence of an anti-repressor, such as a particular sugar, even when the cell contains repressor molecules. This means of regulation can be based on the principle of two different mechanisms, the cis-mechanism and the trans-mechanism. In the cis-mechanism, a binding site for the anti-repressor is formed on the DNA close to the binding site for the repressor. After binding of the anti-repressor molecule to this binding site, the binding of the repressor to the DNA is blocked, it can bind an inducer to its binding site and gene expression can take place (c). If an anti-repressor is not present in the cell, the repressor can bind to the DNA and gene expression does not take place (d). In the trans-mechanism, a binding site is formed for the anti-repressor directly on the repressor molecule. Binding of the anti-repressor to the repressor blocks the ability of the repressor to bind to the DNA and the gene expression can progress (e); in the absence of the anti-repressor, the repressor can bind to the DNA and gene expression is blocked (f). In haploid cells, both types of regulation mechanism are equivalent and both can emerge and be fixed in evolution with the same frequency. In contrast, in diploid organisms, the newly formed allele responsible for the cis-mechanism is recessive and thus the new means of regulation can be manifested only when both genes in the cell carry a binding site for the anti-repressor in their regulation area. In the trans-mechanism, this new feature acts as dominant if the affinity of the anti-repressor – repressor complex for the DNA is greater than the affinity of the repressor for the DNA and the presence of at least one allele for the modified repressor ensures expression of both copies of the regulated gene even in the presence of the repressor. The fixation of advantageous dominant alleles is far more probable than the fixation of similarly advantageous recessive alleles – see the Haldane’s sieve principle (II.4.1.1).
If modification of a certain regulation mechanism were to occur in evolution, for example if it were to begin to be advantageous for the organism not to switch on transcription of a certain gene in the presence of an antirepressor, e.g. a certain sugar, in the environment and gene transcription were to be regulated by a repressor through a cis-mechanism, the change in the means of regulation would require the formation of similar mutations in the regulation areas of both alleles of the given gene.However, this is an extremely improbable event.Gene regulation whose transcription is controlled by a trans-mechanism could develop more readily in such a situation.A single mutation in the gene for the regulation protein, in this case the repressor, would lead to altered regulation of transcription of both copies of the relevant gene simultaneously.In fact, the situation is even more complicated as the regulator gene is located in two copies in the diploid genome and it is not probable that both copies would mutate simultaneously.In order for the above scheme to be valid, the regulator gene must act dominantly, i.e. the new variant of the product of the mutated regulator gene must be capable of ensuring a new type of regulation even in the presence of the products of the original allele of the regulator gene.
It thus follows that, compared to haploid organisms, a great many routes to creation of regulation systems are closed for diploid organisms.This fact could, paradoxically, represent an evolutionary advantage, preadaptation to the emergence of multicellularity.This is a similar effect as that which can be encountered when learning the Morse code.The fastest way to learn it is to use mnemonics, words that contain alternation of short and long syllables correspond to the sequence of dots and dashes for the individual letters (alpha, BE a good boy, COca COla, DOG did it, ...).However, anyone who learns Morse code in this way will never be able to use it as smoothly and rapidly as someone who ignores the possibility of using mnemonics and learns the codes for the individual letters directly as a sequence of dots and dashes.s
In multicellular organisms, the individual tissues must differentiate in different directions; in other words, a specific type of regulation of the expression of tissue-specific genes must occur in each type of cell.Simple types of regulation mechanisms can also be implemented through a cis-mechanism, by a system of activators and repressors bonding to the regulation areas of the individual genes.However, the relatively simple DNA structure does not permit an excessively complicated regulation system; to be more exact, the scope of the DNA regulation sections would grow to an inacceptable length for more complicated systems.In contrast, if evaluation of the combination of regulation signals occurs at the level of the regulation proteins, the emergence and possible functioning of more complicated regulation mechanisms can be imagined.This is true if only because the protein is composed of 20 various aminoacids and thus forms a much more complicated and plastic structure and can react to external instigations in more diverse ways than DNA with the same molecular weight.As soon as evolution decides on a cis-mechanism for the regulation of a certain gene, there is a drastic reduction in the probability that it could, in the future, change to a potentially better trans-mechanism.The evolutionary potential of a haploid organism that employs a cis-mechanism for regulation of a great many genes is thus smaller than that of a diploid organism that, from the very beginning, must employ a trans-mechanism for regulation of its genes.
The reason why a haploid prokaryote never developed into multicellular organisms can thus consist in the fact that the haploid genome employs a gene regulation mechanism with no future for the regulation of a great many genes.This mechanism is sufficient for ensuring cell differentiation into a few types, known, for example, for blue-green algae and sporulating bacteria.However, it is not adequate for ensuring tissue differentiation into the hundreds of different types of cells in a multicellular organism.If the external conditions require cooperation of a greater number of morphologically or biochemically different types of cells, a prokaryote must form a multi-species community of mutually cooperating individuals, such as that known for a great many soil and aquatic bacteria.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.