XIV.5.2.1 In certain situations, the secondary sex ratio can deviate significantly from a value of one
Such a situation occurs, for example, if the population is not a panmictic population, i.e. a population of individuals mating together quite randomly, but rather a population consisting of smaller sub-populations, within which the individuals cross intensively; however, crossing between two subpopulations is very rare.Mathematical analyses of this model have demonstrated that, if the members of both sexes have the same ability to migrate, a very different value of the secondary sex ratio is very easily established (Frank 1986).
A similar situation is encountered, for example, in organisms in which reproduction and thus local competition for sexual partners occurs amongst closely related individuals, in the ideal case amongst siblings (local mate competition).In this case, it is more advantageous for females if there is only one male amongst all the siblings (and he fertilizes all the females in the litter) (Hamilton 1967).In this way, the female can reduce the two-fold ecological cost of sex (the cost of males) (see XIII.2.2).The greatest number of cases in this category are encountered amongst parasites and parasitoids.The value of the equilibrium secondary sex ratio ![]() can be calculated from the equation
can be calculated from the equation
![]() R*= (1-s)/2,
R*= (1-s)/2,
where s is the fraction of daughters fertilized by their brothers (Hamilton 1967).This equation is frequently used to calculate the frequency of superinfection (infection of an already infected host) for parasitic protozoa (Fig. XIV.6).If the frequency of superinfection is low, most sexual reproduction occurs between related individuals.As a consequence, the sex ratio is established during evolution with a bias towards overproduction of females.The frequency of superinfection and the actual intensity of reproduction with genetically unrelated individuals are very important factors from the standpoint of the development of virulence (see Chap. XIX).
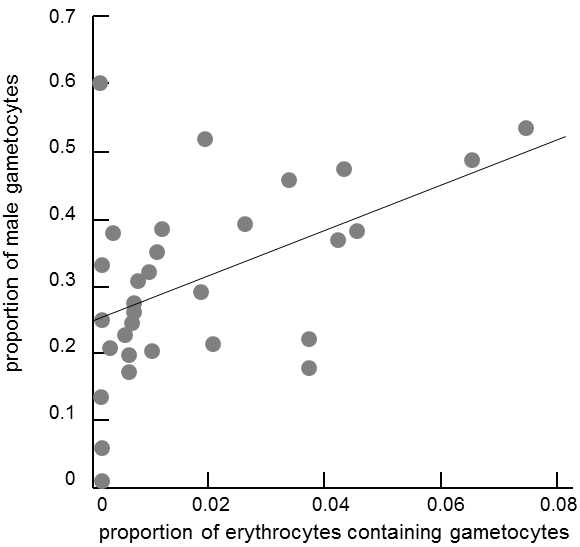
Fig. XIV.6 Shifts in the secondary sex ratio as a consequence of local competition for sexual partners. In some species of parasitic protozoa of the Apicomplexa group, the number of infected red blood cells in the host reflects the number of clones of the protozoa present and thus also the probability of competition for female gametocytes between mutually unrelated male gametocytes. In accordance with Hamilton’s model of local competition for sexual partners, the proportion of male gametocytes increases with the intensity of parasitization (and thus increasing probability of competition between unrelated male gametocytes) in Plasmodium tropiduri (blood parasite of reptiles). Data from (Pickering et al. 2000), modified according to West et al. (2001).
On the other hand, under circumstances where related individuals compete locally for some rare resource (local resource competition) and males migrate more frequently to greater distances from their maternal population, it is more advantageous for a female to produce more males in the progeny (Perrin & Mazalov 2000; Silk 1984).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.