XIX.5.5.1.1 If a parasite is to be transmitted mostly vertically, it must have specific mechanisms in place ensuring its persistence in the host population
By definition, a parasite reduces the fitness of its host.Consequently, genealogical host lines infected by a vertically transmitted parasite are at a disadvantage compared to uninfected lines.As a result, these lines and their parasites should sooner or later disappear from the population.If a vertically transmitted parasite is to survive for a long time in the population, it must have specific mechanisms formed for this purpose.
One of the ways in which a vertically transmitted parasite can ensure survival in the population is by placing those individuals in the host population that are not infected at a disadvantage.Yeast killer factor, dsRNA-viruses transmitted primarily vertically in the population, from parents to offspring, are typical representatives of parasites employing this strategy.The double-helix RNA of these viruses encodes both a toxin excreted by the infected cell into its surroundings and killing yeasts of the same or related species, and also an antitoxin that protects the infected host cell against the action of the toxin.The infected yeast cells expend a major part of their resources for synthesis of the RNA of the killer factor and molecules encoded by this RNA and thus multiply more slowly than their uninfected competitors.Simultaneously, however, in contrast to the unattacked yeasts, they are not killed by the toxin, which, of course, places them at an advantage in intraspecific competition.
Another possibility that is sometimes utilized by parasites is to become indispensible for the host organism.In this case, the parasite does not harm the competitors of the infected host, but “punishes” a host that manages in some way to get rid of the parasite.A number of parasites employ this “drug-dealer strategy”.The restriction-modified systems (RM-elements) of bacteria are a typical example (Fig. XIX.9).(Jeltsch & Pingoud 1996). In this case, the relevant genetic element
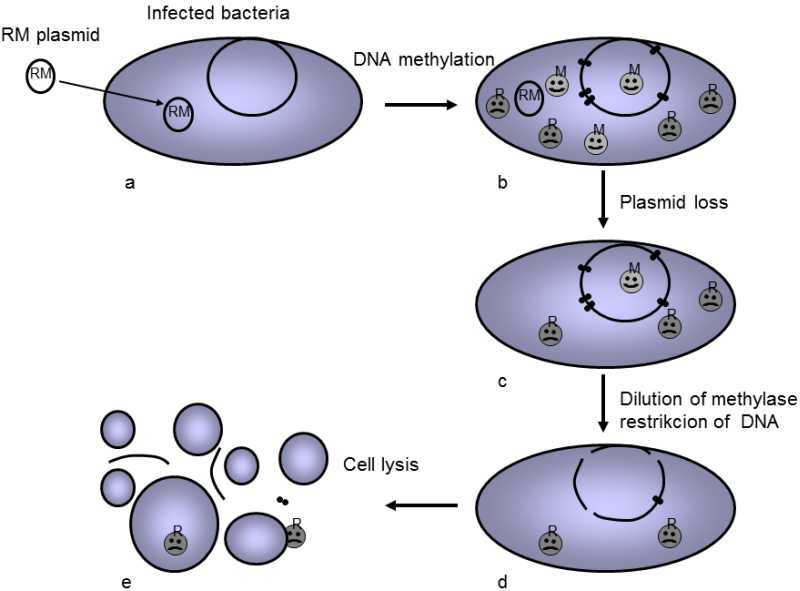
Fig. XIX.9 Restriction-modification system of bacteria as a product of a selfish genetic element employing the drug-dealer strategy. The hypothesis assumes that the restriction-modification (RM) bacteria system did not emerge in evolution as a means of defense by bacteria against their viruses, but as an instrument of a selfish, probably originally extra-chromosomal genetic element, which assisted in maintaining this element in the bacterial population. The model assumes that, in the first step (a) the R-M element penetrates into the bacteria and begins to express the enzyme methylase, which methylates specific target sites in its own and the bacterial DNA. This protects them against cutting by specific restriction endonuclease, which begins to be expressed by it in the next step (b). If any of the bacteria in the population begin to lose this selfish element from this moment on, it stops synthesizing both methylase and restriction endonuclease. Through division of these cells, both the methylase and the endonuclease are gradually diluted in the daughter cells; however, these is less methylase, so it becomes diluted sooner (c). As soon as this happens, the newly synthesized DNA ceases to be protected by methylation (d) and the remaining endonuclease molecules cut it and the bacterial cell dies (e). The selfish element used the drug-dealer effect to maintain itself in the population, i.e. it makes its host vitally dependent on its presence.
(transposone-type genome parasite) both encodes the methylase enzyme methylating a certain short oligonucleotide, for example hexanucleotide, wherever it occurs in the bacterial DNA, and simultaneously also encodes the restriction endonuclease enzyme, which splits the same oligonucleotide if it is not methylated.When the RM-element enters the bacterial cell, it first synthesizes methylase molecules according to it and they then methylate all the relevant sites occurring in the bacterial DNA.After some time, the restriction endonuclease molecules also begin to be synthesized.As, at that time, the endonuclease target sites are already methylated, the bacteria are not harmed by their presence.However, if the bacteria were to get rid of the RM-element some time in the future, both the methylase and the endonuclease molecules would cease to be synthesized.Division of the bacterial cell would gradually lead to a reduction in the concentrations of both enzymes and, as soon as the methylase concentration were reduced sufficiently that at least some of the enzyme target sites would remain unmethylated, the endonuclease would cut the bacterial DNA at these sites and kill the particular cell.This mechanism ensures that rapidly multiplying bacteria that lose this element would not be able to predominate in the population of the particular bacteria in the future.RM-elements were originally considered to be a sort of immune system of the bacteria directed primarily against viruses.However, they do not seem to be particularly advantageous for this function.It is not functional against RNA-phages or against phages with a single-strand DNA-genome, many endonucleases cannot find the relevant sensitive sites in the short phage genomes and, in addition, there is a substantial probability in large bacteria populations that at least some of the phages would acquire the relevant methylation protection prior to splitting and would thus eliminate the relevant defense system of the bacteria in their offspring.Study of some sequences of bacteria and phages has actually shown that palindromes (potential target sites of RM-elements) are eliminated in the genome of bacteria more than in the genome of their phages (Rocha, Danchin, & Viari 2001; Kobayashi 2001).
A great many parasitic bacteria apparently employ a similar strategy, although not in such a drastic form.The existence of the drug-dealer strategy to a certain degree obscures the difference between parasites and mutualists.While symbionts help their host to survive, it somehow caused this dependence of the host on their assistance in the past.For example, it has been observed that, after Amoeba proteus strain D amoebas were infected with rod-shaped bacteria in 1966, their rate of multiplication decreased substantially.However, after several years, the presence of the bacterial endosymbionts ceased to harm the hosts and the amoebas even became dependent on it.When the amoebas were treated with a suitable antibiotic that killed the bacteria, they lost their ability to reproduce (Lee & Corliss 1985).A similar situation apparently also occurred for some helminthes, e.g. filarial, that were infected in the past by bacteria of the Wolbachia genus. It is known that filariasis can be treated with tetracycline, at least in laboratory experiments, where its effect consists in elimination of bacterial symbionts from helminth cells (Dedeine et al. 2001; Stevens, Giordano, & Fialho 2002).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.