XV.4.3 The negative impact of a secondary sexual trait on an individual’s viability can be largely the consequence of sexual selection inertia
An interesting property of sexual selection lies in its marked inertia. At first glance, it might seem that, under conditions where feather length attains dimensions that are disadvantageous for their bearers, sexual selection should “shift to another track”. Females should appear in the population that would begin to select a sexual partner according to other criteria. However, imagine the situation encountered by a female that would begin to prefer males with shorter feathers. Her sons would inherit shorter feathers from their father and will thus have a greater chance of living to reproductive age than their competitors. It might seem that the offspring of the mutated female would have greater fitness. However, this is not true in actual fact. Most of the females in the population will continue to prefer cocks with longer feathers, so that the sons of the mutated female will have reduced sexual attractivness and therefore reducedfitness.
It can be demonstrated that the predominant taste of the females in the flock has a major effect on the reproductive success of males with various phenotypes, especially in polygynic species. In some species of fauna there is even a special ethological mechanism that further strengthens the inertia of sexual selection. It has frequently been observed in birds, mammals and fish (Dugatkin 1996; Agrawal 2001b) that females employ a male’s success with the other females in the flock as one of the most important criteria for selection of a sexual partner (Fig. XV.3).
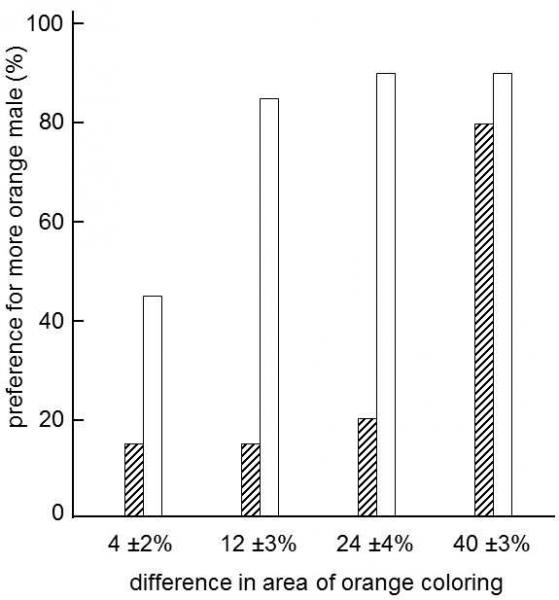
Fig. XV.3 Female preference for preferred guppy males. Under normal circumstances, guppy (Poecilia reticulata) females prefer males with a larger area of orange coloring. In the experiment, females chose between males with different areas of orange coloring on the surface of their bodies under normal circumstances (white column) or after they saw a less orange male courting a different female (hatched column). The results show that, provided the difference in the orange coloring was not very great (about 40 %), females greatly prefer males that attracted the interest of other females, in spite of the smaller area of orange coloring on their body. The height of the columns shows the percentage of cases in twenty independent experiments in which the female preferred the more orange male. Modified according to Dugatkin (1996).
A female who saw that a certain male copulated with a great many other females also frequently prefers this male. This mechanism ensures that the females “need not” rely on their individual taste, but can base their selection of a sexual partner on the predominant taste of females in the particular population. This mechanism is especially advantageous when a female becomes part of a new flock. In order for her sons to exhibit sufficient sex-appeal in the new environment, it is advantageous if the female is able to subject her selection of a sexual partner to the momentary taste of most of the females in the flock.
This model can, of course, not be generalized; in some situations, to the contrary, the effect of the rare male is important. It has been observed in a great many species that above-average success in intrasexual competition is exhibited by males with rare, frequently striking phenotypes (Singh & Sisodia 2000). As was probably first noticed by Daniel Frynta of the Prague Faculty of Science (who, after his custom, did not publish it), the two contradictory effects are, in actual fact, not mutually exclusive and can be active simultaneously in a single population. While most of the females in a particular population systematically prefer males with the most common phenotype, these males tend to compete with one another. When a male with unusual phenotype appears in the population, he has a chance that he will have great reproductive success amongst some females whose individual taste he satisfies. Although most females will prefer the male with the common phenotype, males with unusual phenotype could exhibit the greatest “sexual fitness”.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.