I.14 The direction and general course of biological evolution can be significantly affected by the presence of evolutionary constraints
Another extremely important concept of evolutionary biology, the concept of evolutionary constraints, can be explained on the rather exotic example of flying ears. Evolutionary constraints are generally understood as properties of the structural elements of an organism that constrain the pathways that the evolution of the given species may or may not follow (see also XXVI.7.4). Some biologists, called particularly by their opponents selectionists (functionalists, panselectionalists) are of the opinion that the only constraints that stand in the way of evolution follow from external constraints, i.e. from the laws of mathematics, physics and chemistry. If the existence of a certain structure is not excluded by the laws of the surrounding nonliving nature, then this structure (e.g. flying ears) must be formed in evolution through the effects of the relevant selection pressure. In other words, what is suitable and functional from an evolutionary standpoint and is not prohibited by natural laws will be formed sooner or later.
Other biologists, frequently designated as structuralists, on the other hand, think that certain mutations and thus certain structures can never be formed as, objectively, there exist certain internal constraints, barriers that evolution cannot overcome. In the extreme case, they state that the direction of macroevolutionary development is determined by just these evolutionary constraints, which decide which genetic changes will occur in the particular species. In macroevolution, they attribute a secondary and passive role to selection; according to them, it cannot form new evolutionary forms, but can only constrain the fixation of new forms that are unsuitable from the standpoint of survival of the organism or can hinder this. According to selectionists, flying ears do not exist because they are unsuitable from an evolutionary standpoint. From an aerodynamic point of view, flying organs located on the head would not work effectively; the neck would not be able to support the weight of the body, etc. According to structuralists, for example, flying ears do not exist because, in existing kinds of organisms, a mutation cannot occur that would be capable of modification during embryogenesis in a manner such that sufficiently large bones would be formed in the ear lobes, to which sufficiently strong flying muscles could be attached. Structuralists do, of course, admit the very important role of external limitations following from the physical and chemical properties of the matter from which the bodies of organisms and their environment are formed. The well-known author D´Arcy Wentworth Thompson (Thompson 1942) demonstrated that simple physical forces and geometric laws of growth can substantially predetermine the shapes of individual species of organisms and their organs (Fig. I.16).
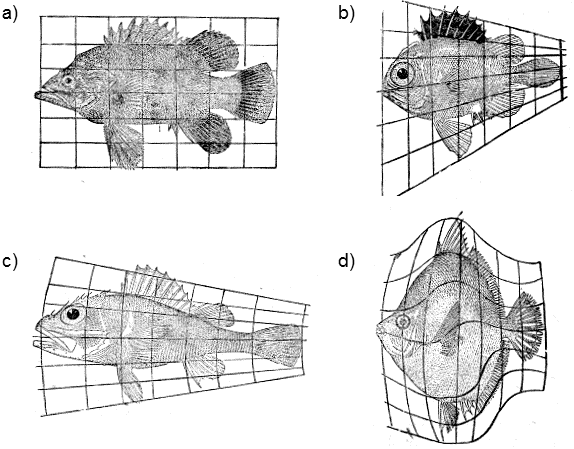
Fig. I.16 Arcy W. Thompson‘s morphological transformations. The shapes of the individual species of organisms are apparently substantially determined by the physical and topological laws applicable during the growth of an individual. The shapes of the individual species can frequently be derived from the basic shape through a simple transformation of the network of coordinates. Fish of the order Acanthopterygia can be considered to have a basic shape close to the morphology of representatives of the Polyprion genus (a), where this basic shape can be obtained by the indicated transformations of the coordinate network of shapes corresponding to the genera Pristigenys (b), Scorpaena (c) and Antigonia (d). Each of these genera is currently assigned to an independent family. According to Thompson (1942).
However, according to structuralists, another important category of evolutionary constraints also consists in the (evolutionarily) historical internal constraints, specifically ontogenetic constraints.These constraints determine which structures can or cannot be formed in the context of the existing ontogenesis. For example, if a certain pattern on butterfly wings is formed by the diffusion of a morphogen from a single place and if, after a certain time, individual dyes begin to be formed at places with certain concentrations, it is clear that only concentric patterns can be formed on the wings and not patterns of a different type (Beldade & Brakefield 2002). It is apparent that, if a certain structure cannot be formed in ontogenesis, it can also not become fixed during the course of evolution. The existence of ontogenetic constraints can thus predetermine the extent of variability in a particular attribute within a species and between species. The possibility that, within a certain taxon, species with a particular type of pattern will predominate and, on the other hand, species with a different type of pattern will not exist at all will not be caused by the fact that the bearers of a certain pattern would be in any way at a disadvantage in natural selection, but simply by the fact that a certain type of pattern cannot be formed in organisms with the particular ontogenetic mechanism. Ontogenetic constraints also exist objectively at a certain moment. However, their character is the result of the historical process of evolution. If the evolution of a particular taxon had progressed differently in the past, the relevant constraints could be completely different or might not exist at all.
The situation is somewhat complicated by the fact that selectionists also admit the existence of at least certain categories of internal evolutionary constraints. However, they assume that these constraints are not caused by the absence of a certain type of genetic variability, but by the above-mentioned one-way character and “short-sightedness” of evolutionary processes or by the fact that many phenotype attributes have the character of trade-offs – i.e. a structure that gives their bearers and advantage in a certain aspect, but a disadvantage in a different aspect. In addition, they are willing to admit that the probability of some mutations is so low that they cannot occur in real time. Thus, in practice, the differences in the opinions of proponents of the two standpoints need not be particularly sharp and will tend to be related to the degree to which the evolution of specific attributes is determined by selection and to which it is determined by evolutionary constraints.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.