I.3.1 In the course of evolution, complex structures emerge through active or passive natural selection, self-organization or sorting by stability
The complex structures that are encountered at all levels of the microstructure and macrostructure of an organism emerge during evolution through the action of a number of various processes. We generally have a tendency to explain their formation through the action of active natural selection. It is probable that a great many complex structures fulfill or once fulfilled some function that was important from the standpoint of survival of the organism and that they could thus be formed through the gradual action of natural selection. The evolutionarily older structures could have been much simpler and could have performed the particular function worse so that their bearers succumbed in competition to individuals in whom mutation led to the formation of a more complicated structure that was simultaneously more functional. The gradual accumulation of these mutations meant that the particular structure developed into the present-day complicated form.
However, complex structures can be also formed by other mechanisms. Passive natural selection is apparently an important mechanism. This factor does not lead to an increase in the functionality of the structure, but does eliminate the consequences of evolutionary changes (mutations) that would otherwise lead to a worsening of the functionality (Stoltzfus 1999). The mechanism of RNA editing in trypanosoma is given as an example of a system that developed into the present-day complex form through passive natural selection. Trypanasomas belong in the class of kinetoplastida and a very complicated means of managing genetic information exists in many species of this taxon (Benne 1992). There are a great many genes of their modified mitochondria that are stored in the DNA in a sort of coded form and must be edited at the RNA level in a very complicated manner before being translated to the relevant proteins. On the basis of the instructions contained in the guide RNA (gRNA), the enzymatic apparatus inserts the individual nucleotides in many sites on the immature messenger RNA (mRNA) and, on the other hand, removes others from the mRNA. In this process, a great many gRNAs specific for the particular gene and coded at various sites in the trypanosome genome gradually participate in a certain order in the process of editing one mRNA. It is very probable that the entire complicated and energetically demanding system does not have any functional importance for trypanosomes and that its formation was the result of passive natural selection. A probable scenario for its formation encompasses gradual multi-stage accumulation of mutations in the relevant genes coding proteins (Fig. I.4). If a line of organisms with this mutation was to survive, it either had to repair the relevant mutation through a reverse mutation or repair its consequences through changes in a different site in the genome. Just such a change could be the formation of an enzyme capable of editing the RNA back to the original form. As soon as such a system formed, even in a very simplified form, the way was immediately opened for its further development. Individuals with a mutation in the particular gene or in genes with similar primary structure ceased to be at a disadvantage in the competition with the unmutated individuals and began to accumulate in the population. The already existing molecular apparatus could be relatively simply modified through mutations so that it was also capable of editing new mutations. The evolution of the editing apparatus was one-directional, genes with a greater number of mutations could no longer be repaired by reverse mutations (because of low probability of co-occurrence of several mutations in the same gene) and trypanosomes became completely dependent on the existence of their editing apparatus.
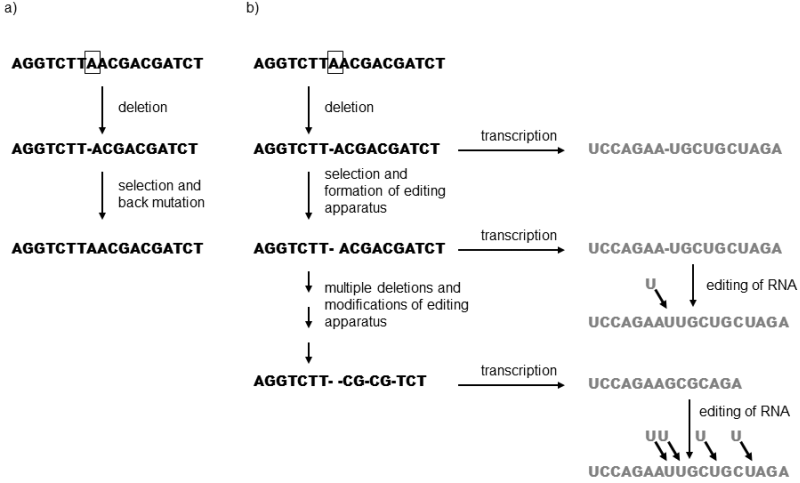
Fig. I.4 Model of the formation of RNA-editing by passive selection. If a mutation occurs in the DNA, for example deletion of one of the nucleotides, which reduces the biological fitness of its bearer, selection pressure begins to act on the mutant and its progeny. As a consequence, over time either revertants to the original form (a) or mutants that are capable of compensating the negative manifestations of the mutation in some way will predominate in the progeny of the mutant. One of the possibilities consists in the creation of an editing apparatus that is capable of repairing the relevant change at the level of the mRNA (b). As soon as one such apparatus is formed, it can be further modified and used to compensate an increasingly broad range of mutations and its universal complexity will gradually increase. After a certain time, the editing process will become absolutely essential for its bearer. The means of additional repairing of mRNA is especially effective and thus highly probable for genes that are located in the cell in many identical copies, i.e. thus for the genes of organelle DNA.
Another factor that could be responsible for the existence of complex structures in living and nonliving systems consists in the sorting of systems from the standpoint of their stability. If certain structures continually emerge and disappear in any system then, at any particular instant, we will probably most often encounter those that emerge most commonly or those that disappear most slowly. While the first category tends to consist of simple structures, the second category may, but need not necessarily, contain more complicated structures. The results of sorting from the standpoint of stability, i.e. gradual accumulation of complex structures, can have some of the features of evolution (Fig. I.5).
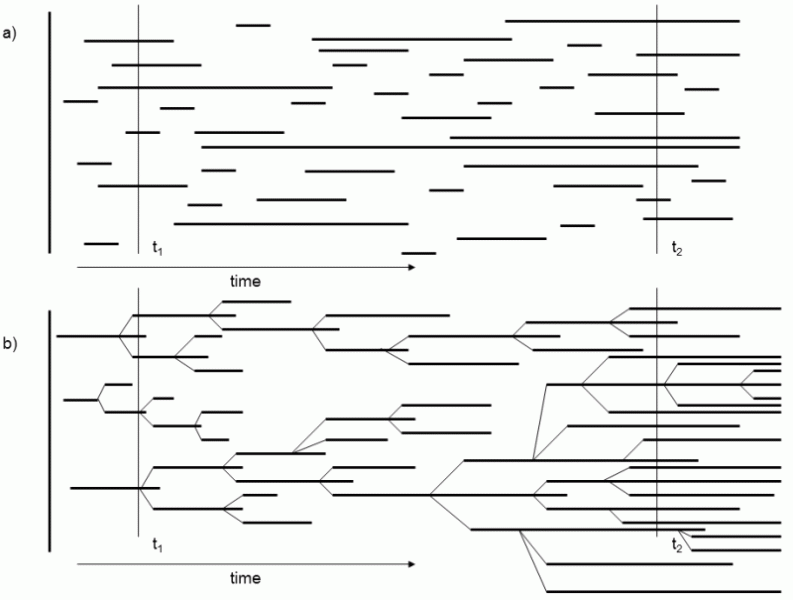
Fig. I.5 Sorting and selection from the standpoint of stability. In case of sorting (a), new mutually independent individuals emerge in the population, where the properties of the newly formed individuals do not in any way depend on the properties of the individuals that exist in the population at the given instant. In contrast, in case of selection (b), new individuals are formed only from parent individuals, from which they inherit their properties, including their probable life expectancy. However, in both cases, the proportion of long-lived individuals increases over time in the population. In case of sorting, this occurs because more long-lived individuals than short-lived individuals always survive to a later stage in evolution of the system. In the case of selection, long-lived individuals reproduce during their lives more times on an average than short-lived individuals and thus leave behind more (once again long-lived) progeny. In case of sorting, the distribution of the life expectancy of newly formed individuals is constant over time, so that the developmental capacity of the system is rather limited. In contrast, in case of selection, the average life expectancy increases constantly over time and the system can constantly develop.
If a sufficiently wide spectrum of various structures is formed in the system, which differ in their resistance to disappearance, more stable structures will gradually accumulate in the system. However, sorting from the standpoint of stability is simultaneously very different from natural selection, as the heredity of the properties of the individual structures is not involved. Structures generally form independently over time and, if they are formed one from another, the daughter structures do not inherit the properties of the parent structures (for an example, see the Life model).
The sorting principle determines the formation of the complex structures that are encountered in the familiar Game of Life, created by the Mathematician John Horton Conway (Gardner 1970), and also the structures encountered in the model of random networks of Boolean logic elements, described in the works of Stuart Kauffman (Kauffman 1993). In the Game of Life, space is conceived in the form of a large chess board.
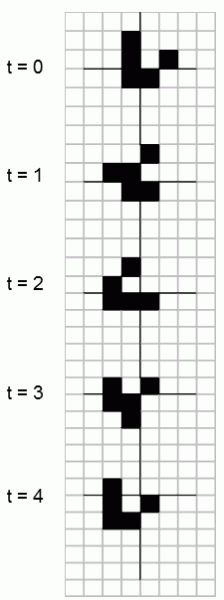
Fig. I.6 Stable shapes are formed in the mathematical Game of Life. Amongst the patterns formed in the Game of Life, various types of “spaceships” are quite remarkable, i.e. stable structures that move across the playing board. A “spaceship” remains stable until it collides with some other shape or until it reaches the edge of the playing board, i.e. it is cyclically converted into several shapes. The figure shows a simple spaceship that, in four steps, always moves one square diagonally downwards to the left.
Each square neighbours on other cells along its sides and in its corners and can be in one of two alternative states, for example black and white. Development of the system occurs in discrete cells according to the following rules: As soon as a white cell is next to three black cells, it becomes black in the next step; as soon as a black cell is next to less than two or more than three black cells, it becomes white in the next step. If a black cell is next to two or three black cells, its state does not change in the next step. On the basis of these simple rules, a system develops from the originally disordered state of randomly distributed black and white cells to a much more ordered state. At various places on the chessboard, relatively large black shapes are formed, which grow or move about (Fig. I.6). Some shapes generate and, on various sides, periodically emit other forms, where the meeting of two forms on the surface can lead to the disappearance of both or only one of them, or several new separate forms are created. The evolution of certain stable structures gradually occurs in the system; their interesting properties have been described in the extensive “life-logic” literature (Neubauer 2003).
The basis for the Kauffman NK models lies in abstract, randomly generated Boolean networks consisting of individual elements capable of transition between two states, on and off (true and untrue). The properties of these elements, i.e. the manner in which they respond to a combination of signals at their inputs, represent the individual functions of Boolean logics. For example, an element of the AND type is converted to the “on” state only if activation signal “turn on” is present at both its inputs, an element of the OR type is converted to the “on” state if the activation signal “turn on” is present on at least one of its inputs, and an element of the XOR type is converted to the “on” state if the “turn on” activation signal is present at just one of its two inputs. The individual networks differ in the number of their elements and the average number of bonds that connect these elements together, i.e. that transfer on-off signals from the outputs of one element to the inputs of another element. If an element is in the “on” state, the “on” signal is present at all its outputs; when it is in the “off” state, the signal “turn off” is present at all its outputs. At the beginning, one of the possible logical functions (e.g. NOT, OR, AND, XOR, etc.) and also a random state, i.e. on or off, is randomly assigned to each element (Fig. I.7).
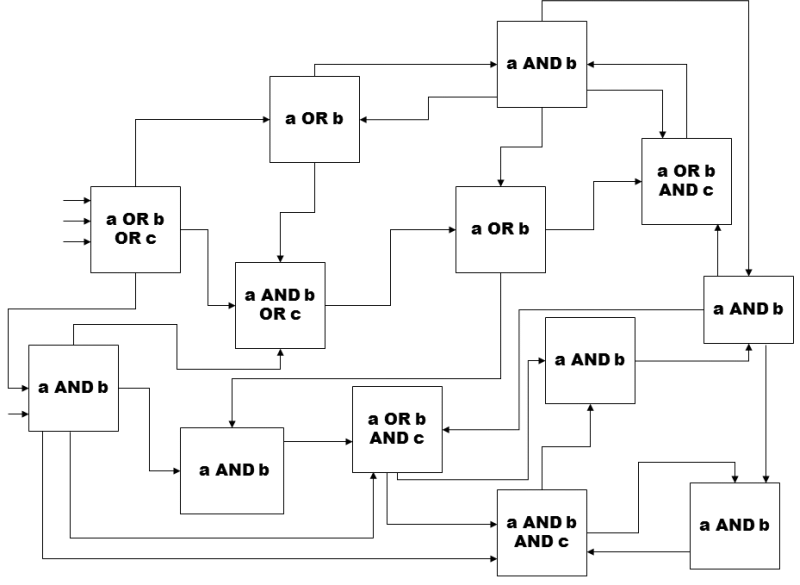
Fig. I.7. Random logical network. The scheme shows a simple logical network containing only elements with two logical operators AND and OR, where each element has a maximum of three inputs. However, Kauffman’s logical networks contained a broader spectrum of various types of logical elements.
The system again gradually develops in discrete steps and, once again, complicated stable or unstable structures are formed in it. Kauffman showed that the system can “freeze”, i.e. stop developing, or pass to a state of chaos, or begin to develop in a direction towards increasing complexity of its structure. He showed that the third, most interesting alternative occurs at the edge of chaos, when there are a medium number of bonds between the individual elements, e.g. an average of two inputs and two outputs, and he simultaneously demonstrated that a great many biological systems occur in this state. (Kauffman 1993).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.