II.1 Genetic information is carried by nucleic acid in contemporary organisms.
Genetic information is a key concept in this chapter. This term will be used to refer to a set of instructions whose main function consists in directing the building of the body of a single-cell or multi-cell organism. Genetic information contributes greatly to directing the building of bodies; this will be discussed in Section II.8. Genetic information (and, to an even greater degree also epigenetic information) is important in directing the maintenance of already created bodies and, in a great many cases, also in controlling biochemical and physiological processes. Genetic information also affects the course of other processes occurring in living systems, including the behaviour of individual organisms. However, genetic information affects these processes very indirectly, mostly through the influence on biochemical and physiological processes occurring in a certain situation in the bodies of organisms. Thus, we can quite easily imagine, especially in multi-cellular organisms, that genetic information is used to create the body of the organism, i.e. is a sort of automatic robot whose behaviour is further determined by autonomous systems, for example the nervous system, whose functioning is more or less not affected by genetic information.
Nucleic acid, in the vast majority of cases DNA, is the material carrier of genetic information in contemporary organisms. The group of viruses, in which genetic information is stored in the form of RNA, probably originated in evolution only secondarily. Genetic information is encoded in DNA in aperiodic sequences of four kinds of nucleotides, i.e. deoxyadenosin-5´-phosphate (A), deoxythimidin-5´-phosphate (T), deoxyguanosin-5´-phosphate (G) and deoxycitidin-5´-phosphate (C), in long, unbranched chains of the double-helix nucleic acid. The two chains are complementary, i.e. the nucleotide containing the base adenine in one chain corresponds to the nucleotide containing the base thymine in the second chain and the nucleotide containing the base cytosine in
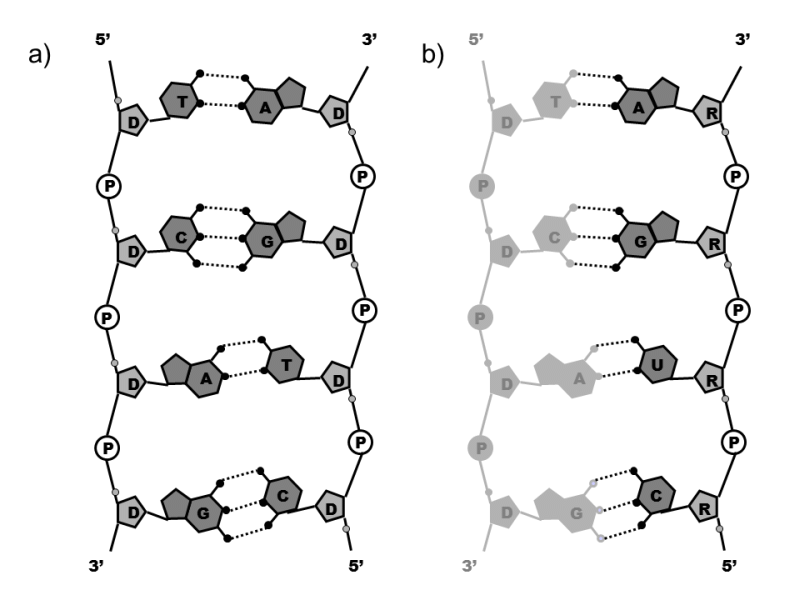
Fig. II.1 The structure of DNA and RNA. The basic unit of DNA (a) is the nucleoside: deoxyribose (D), on whose first carbon atom is one of four kinds of nitrogenous bases bonded by an N-glycoside bond, i.e. two pyrimidine bases – thymine (T) and cytosine (C) – and two purine bases – adenine (A) and guanine (G). Two neighbouring nucleosides are bonded between the 5’- and 3’-carbon atoms, both by a deoxyribose phosphodiester bond (P). Together with the phosphate group, the nucleoside forms the nucleotide. The DNA molecule is thus formed by two linear chains in which four kinds of nucleotides alternate irregularly, i.e. deoxyadenosin-5´-phosphate (A), deoxythimidin-5´-phosphate (T), deoxyguanosin-5´-phosphate (G) and deoxycitidin-5´-phosphate (C). The two chains are interconnected by hydrogen bridges between pairs of complementary bases, AT (two bridges) and GC (three bridges). The chains are mutually anti-parallel, i.e. the 5’ end of one pairs with the 3’ end of the other and forms the familiar double helix structure in space. The structure of RNA (b) is quite similar to the structure of DNA. However, nucleosides are formed by ribose sugar (R) instead of deoxyribose sugar and the pyrimidine base thymine is replaced in the relevant nucleotide by the base uracil (U). Figure b) depicts the RNA molecule at the moment of synthesis, when its base is paired with the complementary base of the relevant DNA section. However, during synthesis, the RNA molecule separates from its DNA template and is thus present in the cell in its single-chain form.
one chain corresponds to the nucleotide containing the base guanine in the second chain (Fig. II.1). Because of this unambiguous allocation of complementary bases, the two chains of the double-helix DNA contain the same genetic information. During the cell cycle, the DNA reproduces by mechanical copying in the replication process. In replication, the double-helix DNA molecule temporarily locally separates into two single-chain molecules, where each of these chains acts as a template for the synthesis of the second, complementary chain. In this way, two double-helix molecules with the same structure, bearing the same genetic information, are formed from a single double-helix DNA molecule (Fig. II.2).
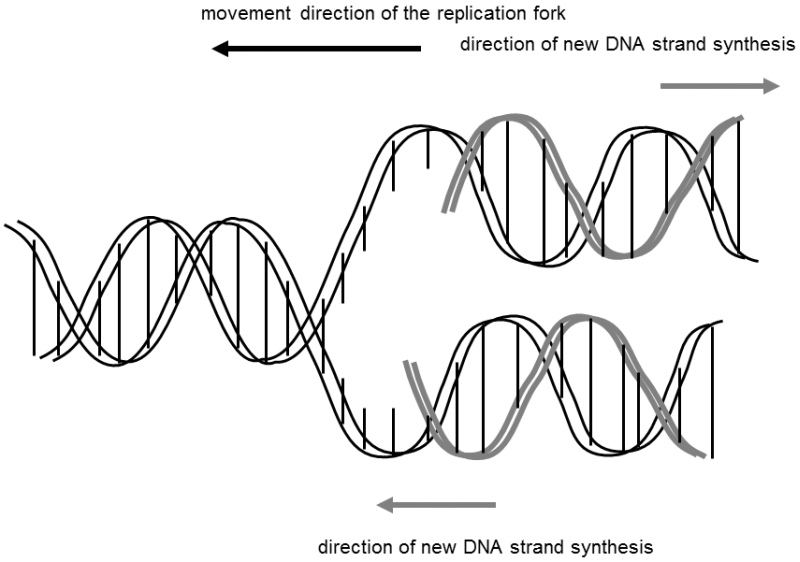
Fig. II.2 DNA replication. During DNA replication, the action of special enzymes causes the two strands of the DNA double helix to be locally unwound and separated and then each strand acts as a template (matrix) for synthesis of the new DNA strand. As the two chains are anti-parallel and the synthesis of a new DNA chain can take place only in the direction from the 5’- to the 3’-end, one of the chains must be synthesized in short sections and these short sections, called Okazaki fragments, are only then connected into a single continuous chain. Replication of the short genomes of viruses and bacteria frequently begins at one point, while the replication of long DNA molecules, encountered, e.g., for all eukaryotes, begins at many sites simultaneously.
All the DNA located in a cell forms its genome. In eukaryotic cells, the genome consists of several parts with different importance. The largest part of the genome is located in the nucleus of the cell and forms the nuclear genome. Other parts are located in the mitochondria (mitochondiral genome)
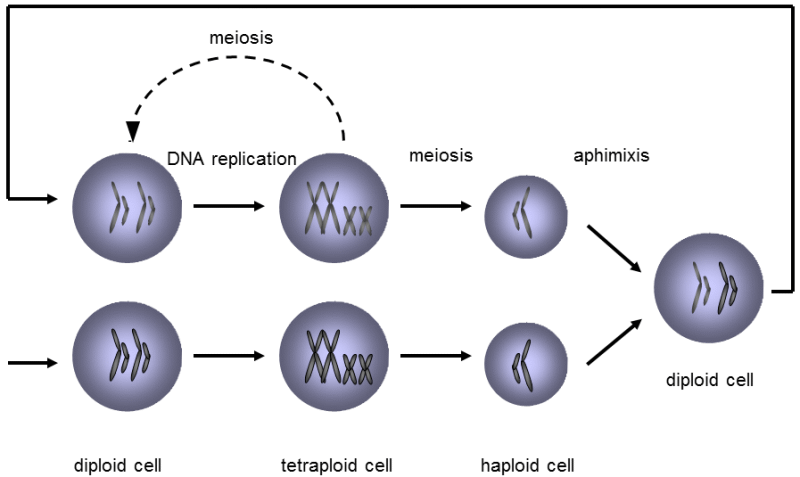
Fig. II.3 Changes in ploidy during the life cycle of a typical eucaryotic organism. In diploid organisms, DNA replication occurs first, so that the cell basically becomes tetraploid from the standpoint of its genetic content. Chromosomes with replicated DNA are formed by two chromatids mutually connected in the area of the centromeres, where the DNA is replicated at the very end. The tetraploid cell can split into two diploid cells during mitosis or into four haploid sex cells during meiosis. The combination of two haploid sex cells, usually derived from two different individuals, i.e. amphimixis, again leads to the formation of a diploid cell.
or in the plastids (plastid genome). Within one part of the genome, e.g. within the nuclear DNA, the genetic information is stored on several (frequently many) independent carriers, on the individual chromosomes. Each chromosome contains one long (double-helix) DNA molecule and a number of other molecules, especially proteins, with structural or regulation functions. In some organisms, the DNA molecule is closed to form a ring, while it is linear in others. A great many eukaryotic organisms are diploid, i.e. their body (somatic) cells primarily contain two copies of each chromosome in their nucleus, i.e. a pair of homologous chromosomes. During the lifetime of the organism, the numbers of chromosome sets can increase in the individual types of cells, or the individual chromosomes and the individual chromosome sets can, on the other hand, be eliminated. Mature sex cells are haploid as, during special nuclear division, reduction division of the meiosis, the number of chromosome sets is reduced from two to one in them. Diploidization of the nuclear chromosome set occurs in the typical case by aphimixis, i.e. the union of two sex cells, mostly originating from two different individuals (Fig. II.3) or by replication of DNA without subsequent cell division. Meiosis also entails two evolutionarily very important processes, recombination and segregation. Recombination is a process involving crossing-over, i.e. exchange of the individual mutually corresponding sections of DNA between homologous chromosomes – the sections on the chromosome derived from one parent exchange places with the corresponding sections of the chromosome derived from the other parent (Fig. II.4). Segregation refers to random separation of one of each pair of homologous chromosome into the nuclei of the forming sex cells (Fig. II.5).
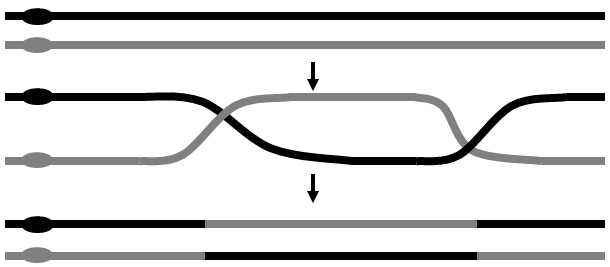
Fig. II.4 Recombination. Recombination entails the exchange of a DNA section between two chromosomes or between two chromatids of a single chromosome. The actual mechanism of recombination is, in fact, far more complicated than indicated by the figure and includes, amongst other things, also synthesis of a new DNA chain and degradation of parts of the older chains. Recombination takes place primarily during meoisis and, less frequently, also during mitosis.
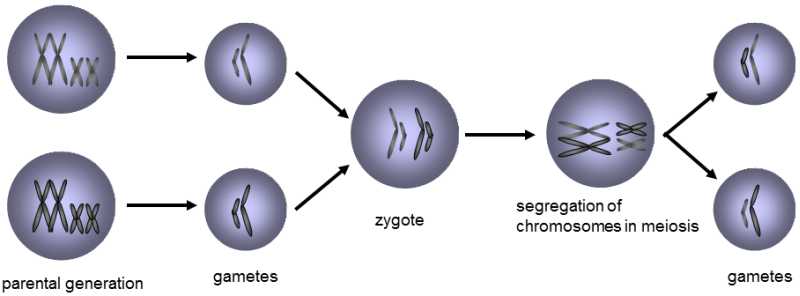
Fig. II.5 Segregation of chromosomes during meiosis. The zygote, i.e. the cell formed by fusion of two parent gametes, bears two sets of chromosomes from the two parents. Random separation of chromosomes into the daughter cells occurs during nuclear division, mitosis and meiosis, so that each of these cells contains a unique genotype formed by mixing the chromosomes derived from the two parents. Simultaneously, the mechanism of nuclear division ensures that each of the cells obtains a complete set of chromosomes, i.e. one chromosome of each type of chromosome.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.