IX.2 The substitution rate of neutral mutations does not depend on population size; the share of selectively neutral mutations in total mutations, and thus also the overall rate, is, however, higher in small populations
The frequency with which mutations are fixed in a certain position or in a given DNA section per time unit in evolution is called the substitution rate.This rate is generally expressed as the number of fixed mutations in the given position per year.The substitution rate must not be confused with the mutation rate; however, it can be concluded that these two rates are numerically equal for neutral mutations (see V.3.3).The mutation rate, i.e. the number of mutations occurring in the given position per time unit for all the members of the population, depends primarily on the accuracy of replication, the efficiency of reparation processes, the intensity of the action of mutagens and the mutability of the sequential motif in the given position of the DNA chain.In contrast, the substitution rate depends not only on the mutation rate at the given site, but also on the intensity and direction of selection that act on the mutation in the given position and in its vicinity.Simultaneously, the substitution rate for selectively neutral mutations does not depend on the size of the population (see V.3.3) (which suggests that genetic drift rather than genetic draft drives the DNA evolution in the studied populations).With growing population size, the number of newly formed mutations in a given position in the population increases linearly, i.e. the mutation rate linearly increases; however, simultaneously, there is a linear decrease in the probability that the newly formed mutation will be fixed by genetic drift.
It must be, however, emphasized that the percentage of mutations that fall in the category of selectively neutral does depend on the size of the population.It is not possible to draw a sharp line between selectively significant and selectively neutral mutations.Basically, only a minimum of them has a selection coefficient equal to zero; most mutations have a negative or positive selection coefficient.It is generally accepted that those mutations whose absolute selection coefficient value is less than 1/Ne, where Ne is the effective size of the given population, act as effectively neutral in the given population.This means that a greater percentage of mutations fall in the category of effectively neutral mutations in a small population than in a large population.As most mutations have a negative selection coefficient and only a minority have a positive selection coefficient and because the probability of fixation of negative mutations is substantially smaller than the probability of fixation of positive or selectively neutral mutations, the total number of mutations fixed by drift over a time unit, i.e. the substitution rate, is greater in a small population than in a large population (see also V.5).
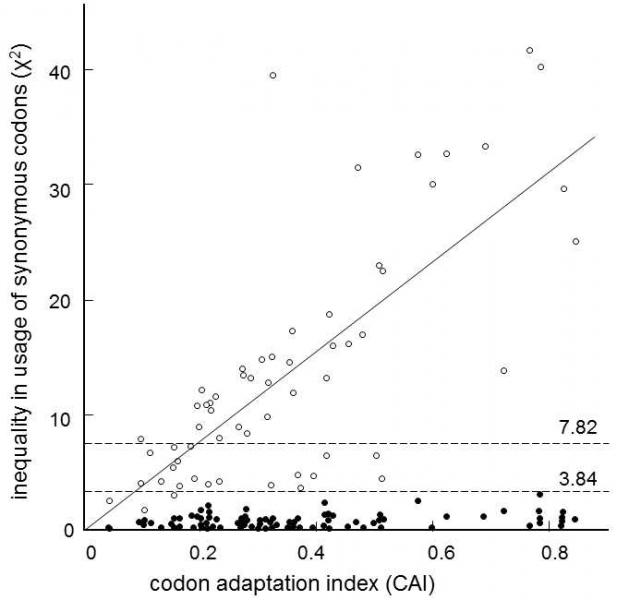
Fig. IX.1. Uneven use of the individual synonymous codons in the bacteria E. coli and Buchnera. The uneven the use of the individual synonymous codons, i.e. codon bias (ordinate) is expressed by value χ2, calculated by comparison with the frequencies of use of the individual codons expected on the basis of the contents of the individual nucleotides. Index CAI (Codon Adaptation Index) (abscissa) reflects the preferential use of the individual synonymous codons in a particular gene in comparison with preferential use of these codons in all the proteins of a studied species. In organisms, CAI is mostly correlated with the degree of expression of the relevant gene. It is apparent from the graph that most of the proteins of the bacteria E. coli (empty circles) exhibit substantially greater unevenness in use of the individual synonymous codons. The value of χ2 exceeded 7.82, i.e. the limit for 5% statistical significance for 274 proteins of 528. In contrast, endosymbiotic bacteria of the genus Buchnera (black circles) employ the synonymous codons much more evenly, so that only 23 proteins of 772 exceeded the value of 5% statistical significance, i.e. 3.84. In addition, correlation between the unevenness in the use of synonymous codons and CAI is completely lacking for endosymbiotic bacteria. Results indicate that, for the genus Buchnera, whose members form relatively small populations in nature that are, in addition, regularly exposed to the bottle-neck effect, i.e. a drastic reduction in population size, the effectiveness of selection is reduced so severely that it is not possible to prevent massive accumulation of slightly negative synonymous mutations, leading to the use of disadvantageous synonymous codons, i.e. codons translated by rare tRNA. According to Wernegreen and Moran (1999).
Comparison with the situation in natural populations has actually shown that relatively the greatest amount of nonsynonymous molecular polymorphism is exhibited by species that tend to form small populations, both for endosymbiotic bacteria in insects (Abbot & Moran 2002)and island species and bird populations (Johnson & Seger 2001).In case of vertically (i.e. from parents to progeny) transmitted endosymbiotic bacteria of the genus Buchnera, it was found that almost all the studied genes developed basically at a faster rate than similar genes in typically horizontally (between unrelated individuals in the population) transmitted bacteria, such as E. coli.Simultaneously, there was a much greater fraction of nonsynonymous nucleotide substitutions amongst the mutations in Buchnera than in E. coli.In contrast to other organisms, there was practically no correlation between the degree of expression of a certain gene and the inequality in the use of the individual synonymous codons in the genes of endosymbionts (Fig. IX.1).All these results demonstrate that a large fraction of mutations fall in the category of neutral mutations in relatively small populations of these endosymbiontic bacteria, which are, in addition, periodically exposed to a short-term reduction in numbers, i.e. passage through a bottleneck, during reproduction of their host.
Similar results were also obtained in studies of ducks (Johnson & Seger 2001).Here it was also found that island species of ducks, forming small populations, have relatively larger numbers of nonsynonymous substitutions in their mitochondrial genes than more numerous mainland species.Of course, there is a greater probability that harm will be caused to the carriers of nonsynonymous mutations than for synonymous mutations.Consequently, only a small number of nonsynonymous mutations are fixed on the mainland, while this number is substantially greater in small island populations.s
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.