V.2.1 The number of homozygotes increases after a large population splits into a number of smaller ones.
Genetic polymorphismof a great many genes exists in every population.The existence of polymorphism is frequently advantageous for the population and species and a great many species have created various mechanisms preventing its loss.There are very many mechanisms preventing relative crossing that, for various reasons, mostly leads not only to a reduction in polymorphism, but also to a reduction in the fitness of individual organisms in the population, i.e. to the inbreeding depression effect.Inbreeding depression is apparently the consequence of the effect of reduction in the average degree of heterozygosity in the individual loci and also of the decrease in the heterozygosity -associated rise of the risk of occurrence of recessive negative mutations in the homozygote state.Some mechanisms responsible for the formation of genetic polymorphism and for its long-term maintenance in the population will be described in Chapter VIII.
The frequency of the individual alleles of a certain gene also determines the frequency of the individual combinations of these alleles – the individual genotypes in the population, i.e. the frequency of homozygotes and heterozygotes for the relevant gene.In a large panmictic population, i.e. in a population where any male can cross with any female with the same probability, the Hardy-Weinberg law holds for the frequency of the individual genotypes of the forming zygotes.This law states that the frequencies of the individual genotypes are unambiguously determined by the frequencies of the individual alleles in the given gene pool (see II.6.1).The frequencies of the three possible genotypes (fAA, fAa, faa) can be calculated for a gene occurring only in two alleles A and a with frequencies of p and q:
fAA = p2 , fAa = 2pq a f aa = q2
The overall frequency of the alleles of the individual genes does not change in any way when a large population is divided into several smaller populations.However, small populations are endangered not only by the inbreeding effect, but some of its alleles are fixed much more rapidly through genetic drift.One of the two alleles is fixed at random in each of the smaller populations so that the frequency of the individual alleles in the overall population once again does not change.Then the Hardy-Weinberg law does not hold for the whole population as the frequency of homozygotes in the overall population will be higher and that of heterozygotes will be lower than would correspond to the frequencies of the individual alleles, i.e. the Wahlund effect is active here (Wahlund 1928).This is caused by the fact that fixation of a particular allele will occur in the individual subpopulations so that heterozygotes will not be formed in these subpopulations at all (Fig. V.4).In the extreme case, after a certain time, a situation could occur where one of the alleles is fixed in each subpopulation so that heterozygote individuals would not occur in the overall population.In practice, such a situation would require that gene flow would not occur at all between the individual subpopulations, i.e., migration of individuals from one population to another would not occur or any migrants in the population would not be able to cross with members of the domestic population.This situation will probably not be common in nature, but could occur, e.g., when an originally continuous water reservoir would be divided through a reduction in the water level in a lake.
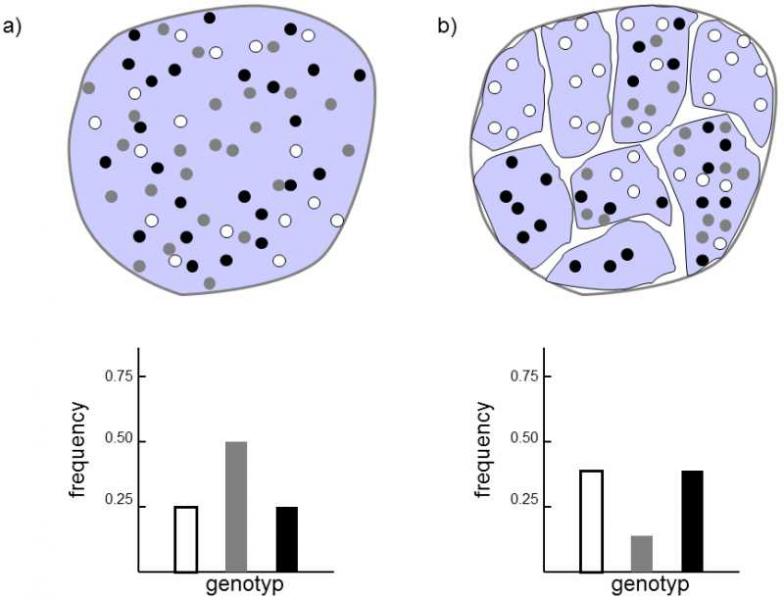
Fig. V.4. Wahlund effect. As soon as the original large panmictic population (a) is divided into a number of smaller mutually genetically isolated populations (b), heterozygotes gradually begin to disappear from the entire metapopulation (denoted as the hatched area). Through the action of genetic drift, certain alleles become fixed in some subpopulations, so that heterozygotes completely disappear from these subpopulations. While the frequencies of the individual genotypes in the original panmictic population and in the individual partial subpopulations correspond to the Hardy-Weinberg law, this is not true of the metapopulation following separation into reproductively isolated subpopulations, as a large part of the heterozygotes is lacking here
The absence of heterozygotes in a real population is, however, frequently a consequence not of spatial fragmentation of the population, but rather the unrecognized presence of two or more cryptic species, i.e. species whose members have extremely similar or even indistinguishable phenotypes but cannot reproduce together.In a great many cases encountered in nature, the absence or a reduction in heterozygotes is caused by other mechanisms that are not related to the action of genetic drift, for example disruptive selection, parthenogenetic reproduction, etc.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.