V.5 Slightly negative (slightly harmful) mutations constitute an important category of mutations.
It followed from equation (5) that the number of mutations fixed per time unit should not depend on the size of the population.However, it is apparent from empirical data obtained from experiments and study of natural populations that more molecular polymorphism is frequently maintained or even fixed over the same time in small populations than in large populations (Nevo et al. 1997).This discrepancy between the theoretical conclusions and fact was explained by the Japanese geneticist T. Ohta (Ohta & Gillespie 1996; Ohta 1998).The core of her explanation consisted in the quite reasonable assumption that most mutations that occur in organisms fall in the category of slightly negative (slightly deleterious) mutations (Fig. V.11).
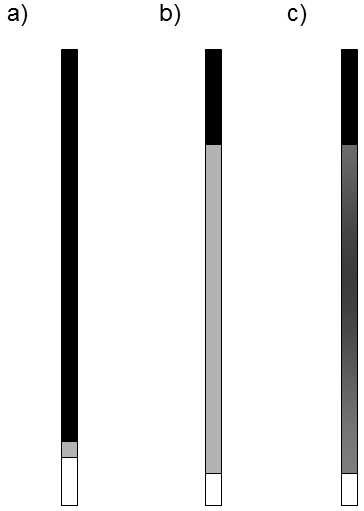
Fig. V.11. Types of mutations. According to the original ideas, the vast majority of mutations are negative (detrimental), a few are positive (useful) and a minimum are selectionally neutral. On the other hand, the theory of neutral evolution assumes that a large portion of mutations is selectively neutral. However, at the present time, it seems that the empirical data correspond most closely to a model of “almost neutral evolution”. This model of evolution assumes that there are very few truly neutral mutations; however, a large portion of mutations act as effectively neutral in small and medium-sized populations and, in large populations, as selectively significant, to be more precise, mostly as slightly negative.
On the basis of study of the mathematical models, W.H. Li derived that mutations that positively or negatively affect the biological fitness of their bearer act as effectively neutral when the absolute value of their relative selection coefficient ![]() Thus, in small populations, the product of the selection coefficient for slightly negative mutations and the effective size of the relevant population (Nes) is less than 1, so that these mutations act as effectively neutral, i.e. their fate is determined by genetic drift and not natural selection.The probability of fixation of these slightly negative mutations is thus approximately equal to their frequency in the population.In contrast, in large populations, the product of the selection coefficient and the size of the population is relatively larger.In a great many cases, it exceeds a value of 1, so that natural selection decides on the fate (elimination) of this mutation.As a consequence, slightly negative mutations are fixed less in a large population than in a small population over the same period of time.In contrast, slightly positive mutations are fixed more in a large population, as natural selection effectively assists in their proliferation here.As there are more negative mutations than positive mutations, it is apparent that overall more mutations are fixed in a small population than in a large population over the same period of time.Thus, the results of theoretical studies are not in any way contradictory to the empirical data:over a given period of time, neutral mutations are fixed with the same effectiveness in large and small populations; however, a greater fraction of all mutations represent effectively neutral mutations in a small population.
Thus, in small populations, the product of the selection coefficient for slightly negative mutations and the effective size of the relevant population (Nes) is less than 1, so that these mutations act as effectively neutral, i.e. their fate is determined by genetic drift and not natural selection.The probability of fixation of these slightly negative mutations is thus approximately equal to their frequency in the population.In contrast, in large populations, the product of the selection coefficient and the size of the population is relatively larger.In a great many cases, it exceeds a value of 1, so that natural selection decides on the fate (elimination) of this mutation.As a consequence, slightly negative mutations are fixed less in a large population than in a small population over the same period of time.In contrast, slightly positive mutations are fixed more in a large population, as natural selection effectively assists in their proliferation here.As there are more negative mutations than positive mutations, it is apparent that overall more mutations are fixed in a small population than in a large population over the same period of time.Thus, the results of theoretical studies are not in any way contradictory to the empirical data:over a given period of time, neutral mutations are fixed with the same effectiveness in large and small populations; however, a greater fraction of all mutations represent effectively neutral mutations in a small population.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.