XII.6.1 The single-cell phase in the life cycle of a multicellular organism is the most important factor limiting the occurrence of intra-individual competition and cell line selection
The most important consequence of the existence of a single-cell phase in the life cycle of organisms is that all the cells in the body of a multicellular organism are mutually genealogically related and thus genetically more similar than the cells occurring in the bodies of two multicellular individuals.Genetic similarity and, if we ignore newly formed somatic mutations, even genetic identity of cells in an organism fundamentally limit the occurrence of mutual competition of cells and cell lines within the multicellular organism – intra-individual competition of cell lines.Here, we will also not consider epigenetic changes and epigenetic heredity of these changes; the reasons and legitimacy of this simplification are discussed in Section II.8.
Intra-individual competition and the consequent intra-individual cell line selection are mostly, but not necessarily always (see XII.6.1.2.1) undesirable from the standpoint of maintenance of the integrity of a multicellular organism.If the cells could compete together, those that were capable of the fastest reproduction and the most effective spreading in the body of the organism would, in time, predominate in the individual.Then these cells would multiply at the expense of those that invest their energy into performing their physiological functions in the body of the multicellular organism rather than into their own reproduction.Those cells that would be capable of preferentially occupying the position and function of precursors of germinal cells would then predominate at the population level.
If the ontogenesis of the organism begins from one cell and amphimixis or some similar process does not occur during its progress, all the cells are genetically very similar.Any phenotype differences can be only of a nongenetic nature, are not passed on to the progeny and cannot become an object of selection (Grosberg & Strathmann 1998).It is significant that the degree of ploidy in the somatic cell line is not reduced during the development of organisms through some form of reduction division.The mitotic cycle occurs through a mechanism including temporary tetraploidization and subsequent separation of the individual chromatids, rather than separation of the individual homologous chromosomes.In the alternative mechanism of nuclear division, i.e. pairing and subsequent segregation of whole two-chromatid chromosomes originally derived from two parent sets, genetically different cells are formed (Fig. XII.4).Competition and selection can subsequently occur between genetically different cells.Somatic recombinations, which occur during mitosis and which can generate genetic variability through the position effect (see XII.6.1.2.1), are also much more common than recombination, which occurs during reduction division in multicellular eukaryotes but not in a great many single-cell eukaryotes (Esposito & Wagstaff 1981).Because recombination certainly plays an important role in the processes of reparation of damaged DNA and somatic cells must manage without this mechanism, it is apparent that there must be a good reason for the absence or rarity of somatic recombination.This reason could be the necessity of maintaining maximum genetic uniformity of the cells within a multicellular organism.
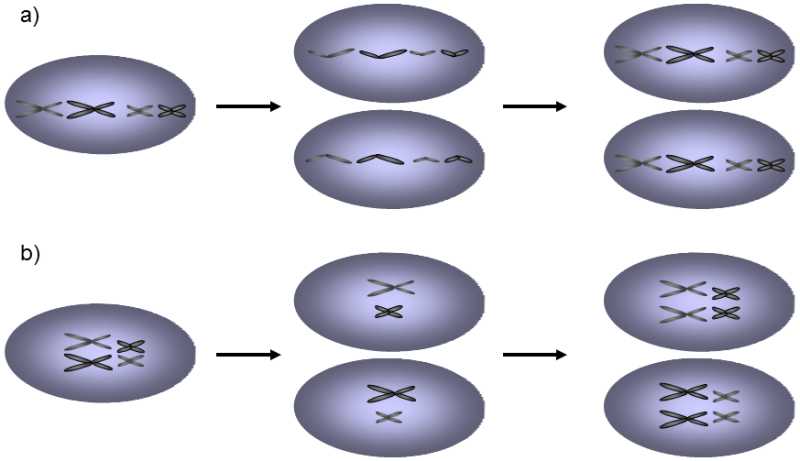
Fig. XII.4 The mechanism of mitosis and genetic variability. In multicellular organisms, mitosis occurs through mechanism (a), in which distribution of the genetic material occurs through separation of two chromatids of each chromosome to opposite fields of the meiotic spindle and subsequently to both daughter cells. If mitosis occurred through mechanism (b), i.e. through pairing of homologous chromosomes and subsequent parting of pairs of these chromosomes to the daughter cells, the daughter cells would acquire a unique genotype as a result of segregation of the chromosomes. The emerging genetic variability would permit selection of the faster reproducing cell line, which could disturb the integrity and functioning of the multicellular organism. The mechanism of classical mitosis can also be expected for single-celled parasitic organisms, as selection occurring within the hosts can lead in parasites to the formation of clones with excessively high virulence, which would kill the host prematurely. To the contrary, the alternative mechanism of mitosis, reminiscent of first reduction division of a meiosis, could occur where it is advantageous for siblings to differ as much as possible, i.e. primarily amongst unicellular organisms
Analogous problems associated with intra-individual selection and analogous means of solution occur in quite different areas than the development of a multicellular organism. These include the area of evolution of the life cycles of parasites and also the area of evolution of eusociality.In the former case, it is, at the very least, striking that a parasite usually reproduces asexually in its host, while the invasive stage, i.e. the stage that passes into the open environment and subsequently to other hosts, almost always originate by sexual reproduction,.Where a parasite reproduces sexually within a single host, the progeny usually pass to other tissues than those in which the parent individual is living.This passage to other tissues is usually connected with irreversible differentiation into another, usually dormant stage of the particular parasite, and is thus also irreversibleThis fact both ensures that genetically different progeny cannot compete with the parent individual and also that a line cannot be selected amongst them that would reproduce to such an degree that it would excessively damage the host and thus reduce the total number of invasive stages that the parasite population produces in the particular host.
The second area is the development of eusociality.It is again, at the very least, remarkable how frequent the formation of eusociality occur in mobile animals forming colonies; i.e. the differentiation of individuals within the colony into casts, frequently even simply into individuals providing for all reproduction and a cast of workers and soldiers.This division of functions once again ensures that the individuals within the colony and thus a sort of superorganism will be as closely related as possible and that competition and gradual selection of individuals preferring their own reproduction over the optimum level of cooperation inside the colony does not occur.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.