XVI.4.1 Altruistic behavior lowers the biological fitness of its bearer and heightens the biological fitness of the individual towards whom it is employed
Altruistic behavior has always been subject to heightened interest from evolutionary biologists. The origin and prolonged endurance of altruistic behavioral patterns cannot be explained by the mechanism of individual (intraspecific) selection; consequently its relatively frequent incidence in nature long remained a mystery to biologists.
Historically, the first suggested mechanism of evolutionary origin of altruistic behavior consisted in group selection. According to this hypothesis, altruistic behavior belongs among traits that were not fixed by individual, but by group selection, because they favor groups (sub-populations), where bearers of these traits appear, at the expense of groups with fewer or no bearers of altruistic behavior.The effect of group selection was long thrown into doubt, mainly because the influence of individual selection in favor of egoists (free riders) inside the groups easily prevails over the influence of interpopulation selection in favor of groups with altruists (see IV.8.2). Today, the opinion prevails that, in a normal (structured) population, consisting of a larger number of continuously emerging and fading sub-populations, this mechanism can be relatively strong and, under some conditions, can even prevail over the influence of individual selection. It can therefore be expected that, in at least some cases, group selection is responsible for the origin and maintenance of altruistic behavior (Alexander & Borgia 1978; Shanahan 1998; Wilson 1975a).
Another mechanism that may be responsible for the origin of altruistic behavior is kin selection (Hamilton 1964a; Hamilton 1964b). As the theory of interallelic selection (the selfish gene theory) stresses, the decisive criterion for fixation or loss of a mutated allele is not how the allele contributes to the fitness of the individual in whose genome it is located, but how it contributes to spreading the copies of the allele in the particular locus inside the gene pool of the population (Dawkins 1976). Some alleles may contribute to spreading their copies within the gene pool so that their bearer increases, through his altruistic behavior, the fitness of the other bearers of the same allele, usually his relatives, at his own expense (Fig. XVI.5). Once again, it seems that the existence of some patterns of altruistic behavior can be explained by this mechanism.
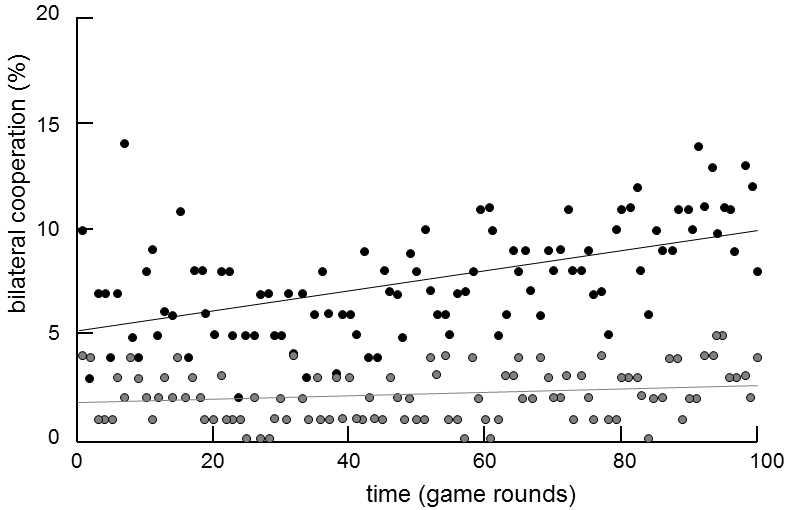
Fig. XVI.5 The impact of genetic relationship on the willingness to cooperate. Willingness to cooperate was tested in 59 monozygotic and 37 dizygotic same-sex twins using the prisoner’s dilemma game. During the experiment, siblings each sat in a different corner of the room and had to raise a red or a blue mark. If both raised blue marks (cooperation), each received 3 pence. If both raised red marks (betrayal), each received one penny. If one raised the blue mark and the other the red mark, the red-mark twin received 5 pence and the blue-mark twin did not receive anything. Each pair of twins played 100 games in total. The chart shows the evolution of average cooperation during 100 games for monozygotic twins (black points) and for dizygotic twins (grey points). Cooperation was expressed as the average of the number of bilateral cooperations (valued at +1) and bilateral betrayals (-1). Unilateral betrayal was calculated as 0 points. The chart demonstrates that the level of cooperation was, on an average, higher and grew continuously during the experiment in genetically more related individuals, i.e. monozygotic twins, while it remained low throughout the experiment in dizygotic twins. After Segal and Hershberger (1999).
Another explanation of the existence of altruistic behavior assumes that it is often actually a case of reciprocal altruism (Trivers 1971; Axelrod & Hamilton 1981). In reciprocal altruism, the individual employs a particular pattern of altruistic behavior only vis-a-vis those members of the population from whom he can, in the future, expect returning of the relevant altruistic behavior. Ethological studies mostly show that individual members of the population constantly “keep a record” of how each member of the population altruistically behaves towards them or towards other members of the population and, according to the degree of his altruism, they behave or don’t behave altruistically towards him.
In species with a sufficiently well-developed neural system and sufficiently well-developed social structure of populations, behavioral patterns have become fixed that lead to punishing less altruistic individuals or even to punishing individuals who do not participate in punishing less altruistic individuals (Gintis, Smith, & Bowles 1901; Okamoto & Matsumura 2000). These behavioral patterns, of course, greatly promote the existence of altruistic behavior. The subject of reciprocal altruism will be further treated in the section devoted to competition of strategies in the sphere of games with repeated interactions between players (XVI.5.3).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.