XVIII.4.1 Races can take place at both interspecific and intraspecific levels
Arms races can also occur at an intraspecific level and this can happen on a relatively short time scale in microevolutionary processes.The members of the male or female sex can increase their fitness at the expense of the members of the other sex.Under normal conditions, the speed of evolution of males and females is comparable, so that it cannot be observed that the members of one sex would gain an apparent advantage over the members of the other sex.However, by suitably arranging an experiment, we can ensure, for example, that the females will not be able to respond to the evolutionary drives of the males.The experiment is performed in that the males are allowed to interact ecologically, ethologically and sexually with the females of a certain strain for a number of successive generations, where only the males of the obtained progeny are allowed to survive.The females are replaced in the experimental population in each generation from the stock population (and are thus completely “naive” from the standpoint of interaction with the particular selected line of males).The results of such an experiment performed on drosophila indicate that the males gained a considerable advantage over the females after 41 generations (Rice 1996).They achieved copulation with the females of the particular strain much faster; following copulation, the time before the females began to copulate with some other male was prolonged and, in case of multiple copulation with their own and foreign males, only a small percentage of the eggs were fertilized by the foreign males.The females lost out in the evolutionary battle, not only in the limited ability to chose, from their point of view, the best father for their progeny, but also in shortening of their lifetimes following copulation with the test males (but not with control males) (Fig. XVIII.2).
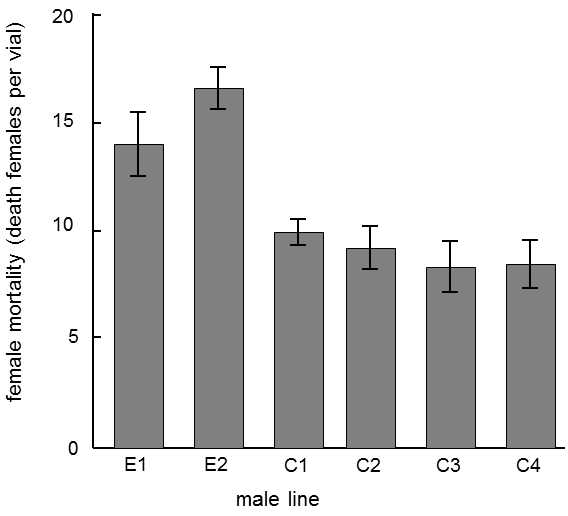
Fig. XVIII.2 Coevolutionary battle between male and female drosophila. In a special experimental arrangement, females were prevented from responding to the evolutionary moves of males for 41 generations (the females in the cultivation vessel were replaced in each generation by “naive” females from the stock culture). As a consequence, the adapted males had greater fitness in reproducing with females of the particular strain compared with the control males (they became the fathers of a substantially larger number of progeny than would correspond to their numbers in the test vessel). However, their fitness increased at the expense of the females. When the females mated with the experimental males, they exhibited greater mortality than when they mated with the control males (C1–C4). According to Rice (1996).
According to some authors, the coevolutionary battle between the sexes in sexually reproducing species is the main reason for the genetic divergence of allopatric populations and is thus indirectly an important motor for allopatric speciation (Martin & Hosken 2003).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.