XXI.3 New species can evolve allopatrically or sympatrically, i.e. without contact or in contact with the parent species
A new species can be most readily formed by gradual evolution outside of direct contact with the parent species, i.e. by allotropic speciation (Fig. XXI.3). If, for example, a geographically isolated
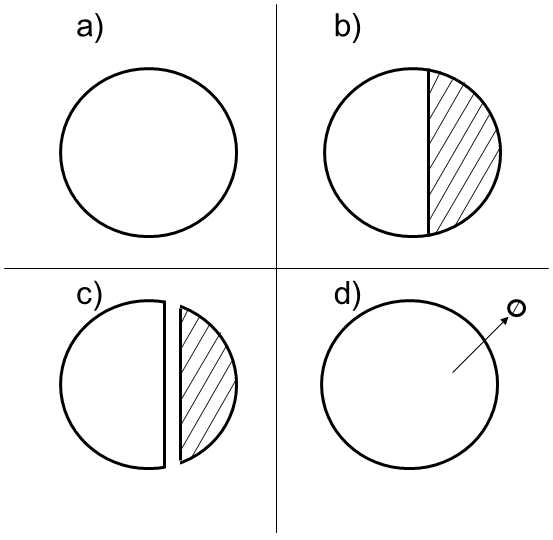
Fig. XXI.3 Classification of geographic speciation. In sympatric speciation (a), the individual species develop within the range of the parent species, where the members of both species are in ecological and genetic contact during speciation, i.e. there are no spatial barriers between them that would prevent their mutual competition and crossing. In parapatric speciation (b), the two species occupy separated ranges at the moment of speciation; these ranges are, however, in contact and gene flow can occur between them through migrants. In allopatric speciation of the vicariant (dichopatric) type (c), the ranges of the two species are separated at the instant of speciation and exchange of migrants does not occur between them. The two ranges were formed by division of the original range by a geographic barrier into two parts of approximately the same size. In peripatric speciation (d), the ranges of the two species are also separated at the instant of speciation and gene flow does not occur between them. However, the range of one species was formed by the separation of only a small part of the population of the parent species, most frequently during colonization of a new territory or survival of a residue of the population in a spatially limited refuge following contraction of the range of occurrence of the original species.
population is formed, which branched off from the population of the original species, and this population is reproductively isolated from the parent population for a sufficiently long time, genetic changes can gradually accumulate in its gene pool that finally lead to phenotype and subsequently also ecological differentiation of the two populations. If the two populations come into contact before they are sufficiently differentiated, the two species can again merge into a single species. Otherwise, the two species can exist sympatrically next to one another (if there was differentiation of their niches) or one of the species can force the other species out of the location or even globally.
Sympatric speciation is the opposite of allopatric speciation. In sympatric speciation, a new species is formed in the same territory as that occupied by the parent species. Occurrence in the same territory at the time when speciation occurs is a necessary but not a sufficient condition for the particular speciation to be considered to be sympatric. If the members of the new and original populations basically do not meet in the particular territory, this would not be sympatric speciation. The main difference is that allopatric speciation is accompanied by the formation of internal reproductive barriers between differentiating species in the presence of already existing reproductive barriers, while sympatric speciation occurs in their absence. For example, if a parasite transmitted by direct contact happens to jump over to a different species of host living in the same territory as its original host, the two populations of parasites will continue to exist sympatrically in their range of occurrence. In actual fact, no interactions need occur between the parasite populations on the original and new host species, specifically because of the minimum of physical contact between the members of the two host species, and thus there will be no exchange of genetic information or competition for resources. The two parasite populations can gradually diverge into independent species. However, these will certainly not be sympatric speciation, as internal reproductive barriers would be formed in this case after the formation of external reproductive barriers. Similar cases, when the species live in the same territory but have island ranges at different places, are mostly called microallopatric. In true sympatric speciation, the members of the two populations must constantly meet during the evolution of the new species.
A large fraction of instant speciation, for example polyploid speciation or hybridization speciation, has the character of sympatric speciation. In these types, one-step formation of reproductive isolation barriers precedes, or even causes, phenotypic and therefore also ecological differentiation of a new species. It is understandably rather questionable whether it makes sense to differentiate sympatric andallopatric speciation in cases of instant speciation. However, sympatric speciation also includes gradual ecological speciation, which occurs through the action of disruptive natural selection or ethological speciation. These gradual speciations are accompanied by the accumulation of changes that eventually gradually lead to complete reproductive separation, where constant gene flow between the populations consistently prevents accumulation of differences. The possibility of formation of a new species through gradual sympatric speciation thus continues to be a matter for discussion.
Basically, parapatric speciation forms an intermediate link between allopatric speciation and sympatric speciation (Gavrilets 2000; Pennisi 2000a). Parapatric speciation could occur, for example, in not very mobile species whose members form mutually neighboring local populations within their range of occurrence. Crossing occurs between the organisms within these populations, while the gene flow between populations is much less and tends to be mediated by isolated migrants. The fact that the individual populations are constantly in contact means that there is a continuous gene flow between their members, even during speciation. This, of course, complicates their phenotype differentiation and the formation of post-zygotic reproductive isolation barriers. However, if the gene flow is sufficiently limited, for example because the ranges of occurrence of the two populations come into contact only at a certain restricted site, the individual populations can nonetheless adapt to the local conditions of their environments. Crosses born as a consequence of penetration of migrants into the territory of a foreign population exhibit a combination of the traits of the two populations and consequently have suboptimal phenotype and lower fitness in both environments. They are thus gradually eliminated from the population, again reducing the gene flow between the two populations.
The effectiveness of parapatric speciation, similar to the effectiveness of gradual sympatric speciation, is sometimes doubted. Mathematical models indicate that even very small gene flow, for example, exchange of a single individual per generation between the two populations, is mostly enough to prevent genetic differentiation of a new species. However, in actual fact, these conclusions are valid only for differentiation occurring through the action of genetic drift. If more effective processes participate in the differentiation of the two populations, such as evolutionary drives or selection, the intensity of the gene flow would have to be much greater to prevent differentiation.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.