XXI.4.2.2.2 Postzygotic reproductive isolation mechanisms include zygote mortality, partial or total hybrid inviability, partial or total hybrid sterility and reduced viability and fertility in the F2-generation due to chromosomal or gene incompatibility
Postzygotic mechanisms represent a very important category of reproductive isolation mechanisms, i.e. mechanisms that are active after fertilization of the oocyte by the microgamete of a foreign species. Compared to prezygotic mechanisms, they have the disadvantage from the viewpoint of the species that they simultaneously reduce the fitness of the reproducing individual. In case of mortality of the zygote, the individual must invest energy and time into the actual act of copulation and production of gametes. The investments into production of imperfect progeny can be even greater for the other types of postzygotic barriers. In the extreme case, the abortive development of hybrid zygotes may even kill the maternal organism. Although the action of normal mutagens leads much more frequently to mutations causing inviability or sterility, in contrast the sterility of hybrids or their progeny is far more common in nature than their inviability (Johnson & Kliman 2002). This is apparently a consequence of the fact that the sterility of hybrids is frequently caused by defects that occur during segregation of chromosomes during meiosis. This incompatibility at the chromosome level evidently often occurs as a consequence of the existence of mutually incompatible changes in the chromosome morphology that were rapidly fixed by evolutionary drive after splitting off from the common ancestor (see VI.3.5.1).
Incompatibility at the chromosome level seems to constitute a very important postzygotic reproduction barrier and its formation as a consequence of chromosome mutation can substantially affect the evolution of a new species. On the other hand, it should be recalled that the reduction in the fertility of hybrids of two species with partly incompatible karyotype cannot, in itself, prevent the flow of the individual genes between two populations or related species. As soon as at least some of the hybrid progeny are fertile, recombination will occur in their genomes. Recombined chromosome with the morphology of species A but bearing genes of type B will subsequently introduce foreign genes into the gene pool of both species.
Incompatibility at the gene levelis another source of incompatibility of foreign genomes. The products of some genes of one species cannot properly cooperate with the products of the genes of the other species, so that they form dysfunctional molecular complexes and, at the level of the organism, dysfunctional organs, in crosses. The Dobzhansky and Muller model describes the evolutionary formation of this gene incompatibility (Muller 1939; Dobzhansky 1936) (Fig. XXI.7). At the
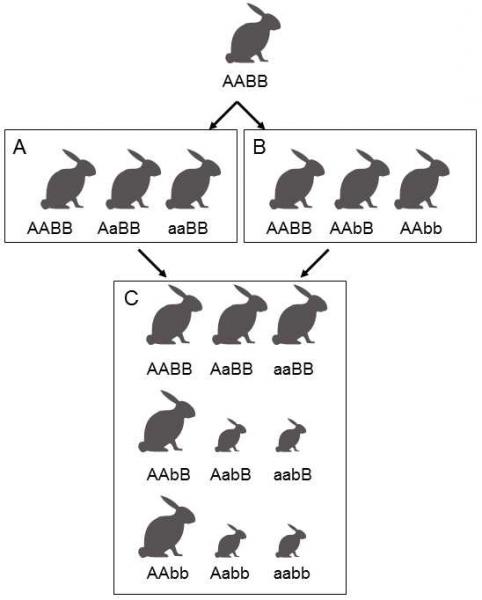
Fig. XXI.7 Dobzhansky and Muller model. The original species had only alleles A and B at both loci. After separation into two genetically isolated populations and subsequent differentiation into two separate species, allele a spread (however was not fixed in our case) in one species and allele b spread in the second species Alleles a and b are mutually incompatible, but do not meet in either of the species, so that their negative effect on the fitness of an individual cannot prevent their spreading and potential fixation. However, if the members of the two species cross, the progeny will contain individuals carrying both new alleles, a and b. These individuals will have lower fitness or will even be unviable. If there are a greater number of similar loci with mutually incompatible alleles in the given pair of species, the vast majority of the progeny will be thus affected.
instant when the gene pools of two populations of one original species are separated (for example, through the effect of spatial isolation), new alleles can be accumulated in the individual loci of one or the other population. The new alleles are always compatible with the alleles originally present in the gene pool of the population (alleles occurring in the same locus and alleles occurring in different loci); otherwise the individuals with the new allele would have reduced fitness and a substantial increase in the frequency or even fixation of the new alleles could not occur. However, the new alleles present in one and the other population need not be mutually compatible, as they never occurred in the same individual during evolution, and their mutual compatibility was thus never tested by natural selection. If the two populations are separated for a sufficiently long time, such a large number of mutually incompatible alleles accumulate in their gene pools that very effective postzygotic reproductive barriers are formed. If the new alleles are evolutionarily fixed in both species, all the hybrids would have approximately the same reduced fertility and viability. In actual fact, a great many new alleles do survive simultaneously in the population together with the old alleles for a long time, most frequently as a substantial component of neutral polymorphism. Consequently, hybrid individuals can differ considerably, even in a single family, in the degree of reduction of their fitness. The existence of postzygotic barriers is thus very often manifested both in reduced fitness of hybrids and in reduced percent of viable progeny formed by hybridization.
Also in the case of genetic incompatibility, functions connected with reproduction, i.e. the fertility of crosses, are usually affected first. This is apparently a result of the fact that the genes participating in these processes undergo very rapid evolution as a result of the co-evolutionary battle between males and females and partly also as a result of intrasexual competition, especially in males, i.e. the effects of sexual selection.
Genetic incompatibility need not necessarily be manifested in the F1-generation, but can appear in later generations. This is a result of the fact that the genome of crosses in the F1-generation contains the full chromosome set of both participating species. Thus, together with dysfunctional molecular complexes, composed of the products of both species, also fully functional complexes, composed of the molecules of one or the other species, can be formed in the cells of crosses. However, if, in further generations, the hybrids cross together, hybrid breakdownoccurs (Fig. XXI.8). The genome of the crosses no longer contains the complete chromosome sets of both species, but rather a set of chromosomes some of whose chromosome pairs are from the first and others from the second species. Only then can some cases of genetic incompatibility be manifested (Davies et al. 1997; Turelli & Orr 2000).
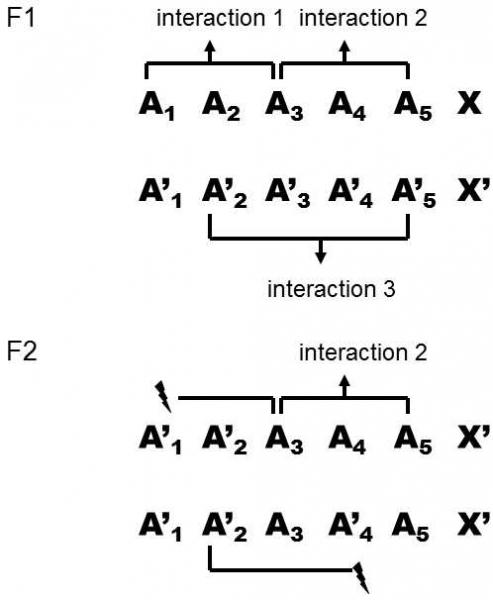
Fig. XXI.8 Hybrid breakdown in the F2-generation. In an F1-generation hybrid, the genotype of the individual contains the complete chromosome sets of both species. Interactions occur between the products of some chromosomes within a single chromosome set, for example they form molecular complexes together; however, the products encoded on the relevant chromosomes of various species cannot interact. In the F2-generation, most progeny acquire some pairs of chromosomes from one species and other pairs from the other species. This means that these F2-individuals do not have a complete chromosome set from either species. Thus some interactions (denoted by “thunderbolts”) cannot occur as some molecular complexes are not formed here or are dysfunctional.
A frequent reason for genome incompatibility consists in inactivation of a nonidentical copy of a gene that multiplied in the genome by gene duplication (Taylor, Van de Peer, & Meyer 2001). Inactivation and subsequent deletion of one of the two copies is the most frequent fate of duplicated genes. In one species, for example, a copy of a gene located on chromosome 1 can be inactivated, while inactivation of a copy of a gene on chromosome 3 occurs in the other species. If the F2-hybrid obtains both copies of the gene on chromosome 1 from the first species and both copies of the gene on chromosome 3 from the other species, it will not have a functional copy of this gene and thus will frequently be unviable.s
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.