XXII.5.3.5 Ecosystems, similar to many other complex systems, can experience spontaneous periodic or aperiodic occurrence and spreading of sudden changes
The mutual relationships between the individual species within an ecosystem form a complicated network of positive and negative relationships.Theoretical analysis of similarly complicated systems has shown that a complicated temporal or spatial structure is frequently formed spontaneously in them, i.e. without input of information from external sources (Kauffman 1993).The existence of temporal structures can be manifested, for example, in that apparently spontaneous disturbances (defects) appear periodically in the system, i.e. sudden changes in the states of a greater number of elements.The energy for these disturbances can be derived from inside the system and thus it can be exhausted after a longer period of time, or can come to the system from the surroundings.The intensity (extent) of the disturbance that occurs in the system at a certain moment is simultaneously not directly dependent on the amount of energy that comes into the system from its surroundings at the given moment, i.e. on the magnitude of the external stimulus.In dependence on the momentary state of the system, a weak stimulus from the surroundings can cause a large disturbance while, at other times, a very strong stimulus can cause only a minimal disturbance.
If the logarithm of the intensity of the disturbance is plotted against the logarithm of the frequency of disturbances of a given intensity, a straight line is very frequently obtained (Fig. XXII.6).In this case,
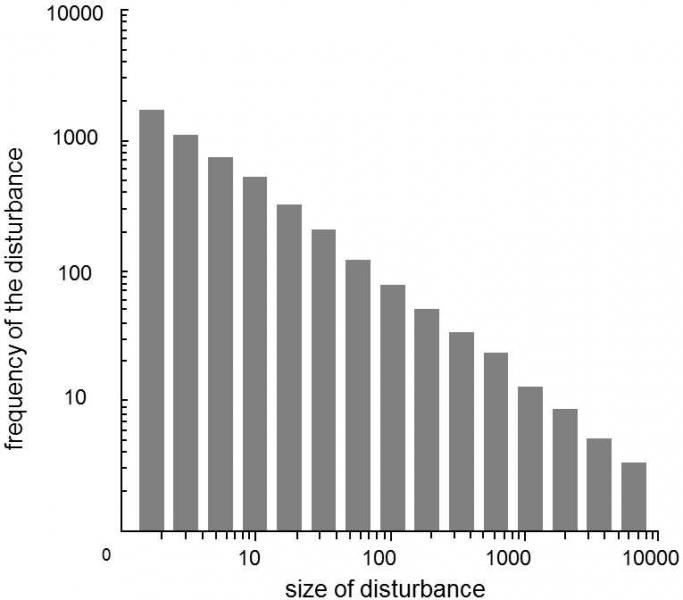
Fig. XXII.6 Increase in the diversity of tetrapods. The graph depicts the progress of the gradual increase in the number of families of terrestrial vertebrates during the Phanerozoic. Modified according to Benton (1997).
we say that a power law governs the distribution of the frequency of disturbances of a certain intensity.This type of distribution of the frequency of disturbances of various intensities differs substantially from normal orPoisson distribution, in which the magnitude of the disturbance would be limited from above, to be more exact the frequency of disturbances exceeding a certain value would be completely negligible.For disturbances respecting a power law, disturbances of low intensities also predominate and the frequencies of larger disturbances gradually decrease; however, their frequency and especially effect on the behavior of the system can certainly not be neglected.A striking property of this type of distribution is that its shape does not depend on the scale employed.If the frequency of disturbances is first measured on a time scale of years and subsequently on a scale of millions of years and we forget to describe the axes in the obtained histograms, it is not possible to distinguish them later on the basis of their shapes.
The described behavior of complicated systems is reminiscent of the evolution of biodiversity occurring on a paleontological scale in a great many respects, including the shape of the distribution of the frequency of disturbances of various magnitudes.The mechanical model of such a system is a pile of long-grain rice, onto the top of which a thin stream of more rice is constantly poured.This phenomenon can be best observed if this pile of rice is formed between two panes of glass or at least against the inner wall of an aquarium.Over time, it can be observed that variously large avalanches of grains run down the surface at irregular intervals, analogous to the waves of extinction.It is not possible to predict when an avalanche will start.However, for each type of rice, it is possible to calculate the characteristic distribution of avalanches of various sizes and, on the basis of this distribution calculated for a shorter period, statistical techniques can be employed to estimate the period before an avalanche of even greater intensity will occur, i.e. an avalanche greater than that which occurred during the original, shorter time interval. This method, which is based on the statistics of extreme values, is employed, for example, by seismologists to estimate the time before an earthquake of a certain intensity will occur, on the basis of seismological records covering a shorter time interval.Apparently not only variously large avalanches of rice and mass extinctions that occurred during the history of life on Earth, but also earthquakes, volcanism, atmospheric phenomena and similar processes also have similar frequency distributions.
Some authors assume that the reason for the similarity in the distribution of mass extinctions and other types of regularly occurring natural processes is that a similar state is characteristic for ecosystems on a long-term scale as that which can be encountered for the mentioned pile of rice, i.e. a state that is mostly termed self-organized criticality (Bak, Tang, & Wiesenfed 1988; Tang & Bak 1988).In general, a region of the state space of a certain system in which there is a sudden change in its behavior, for example from ordered to chaotic, is critical.In the case of self-organized criticality, the system spontaneously remains, i.e. has an attractor, in the vicinity of the critical point or points from which it can suddenly change to two different states, for example to the avalanche – quiescent or mass extinction – background extinction states.Under the influence of minor cumulative changes, e.g. accumulation of grains on the top of a pile or the extinction or emergence of a species, local disequilibrium occurs that, when it exceeds a certain level, is suddenly eliminated by the sliding of an avalanche of grains to the bottom of the pile or the extinction of a greater number of species.If the system is outside of the critical region, the magnitude of the disturbance is limited from above.If it is in the critical region, disturbances can occur with an extent that affects the whole system.
If the changes in the extinction rate over time were actually a consequence of self-organized criticality, this would mean that the intensity of extinction would basically not be related to the intensity of external stimuli and background and mass extinction would have the same cause.Study of mass extinction on the basis of models of self-organized criticality is a favorite pastime of theoretical biologists.Some published models, for example the Kauffman NK-models of Boolean networks, require that the ability for their transition to a state of self-organized criticality be facilitated in advance by suitable choice of parameters – for the NK-models the average number of inputs per element (Kauffman 1993).Understandably, suitably adjusted parameters that make it possible for a system to pass into a state of self-organized criticality can ensure both natural selection and sorting from the standpoint of stability.If systems with the correct number of inputs and outputs per element have greater evolutionary potential than other systems, then it is quite natural that they will be encountered in nature.Other models spontaneously enter a region of self-organized criticality of their state space from almost any arbitrary initial state (Bak & Sneppen 1993; Bak, Tang, & Wiesenfed 1987; Boettcher & Paczuski 1996; Head & Rodgers 1997; Vanderwalle & Ausloos 1995; Vanderwalle & Ausloos 1997).
For the above-described models, the value of the exponent determining the shape of the distribution of the frequency of extinction of various magnitudes in the relevant equation is different from the values measured on the basis of paleontological data; however, models exist that have values of this exponent corresponding to the empirical data (Newman & Roberts 1995; Sole & Manrubia 1996).However, a distribution of the frequency of disturbances corresponding to the power law can also be obtained on the basis of models that do not anticipate the participation of self-organized criticality but which, for example, assume that the source of the individual extinction lies in the interaction of random changes in the external environment with evolutionary processes occurring in the biosphere (Newman 1997).In addition, it is apparent that the behavior of sufficiently complicated models is so highly variable that paleoecological (and also any other) processes can be very readily modeled when the parameters are suitably adjusted.However, with such variable systems it is very difficult to demonstrate that their behavior is actually determined by processes following from the existence of self-organized criticality and that they cannot be explained, for example, by the effect of random disturbances arising from the surrounding environment.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.