XIX.5 Host-parasite coevolution often leads to changes in the pathogenic manifestations of parasitosis
The ability of a parasite to survive and reproduce in the host organism and the ability of the parasite population to survive consistently in the population of the host species are a consequence of gradual evolutionary adaptation of the parasite to the particular host species (Kaltz & Shykoff 1998).If the host species comes into contact with a new range of potential parasites, for example if it invades a new territory, it is generally resistant to parasitization by unspecialized parasites.Escaping from the reach of the original parasites is usually considered to be the most important cause of the ecological success of a great many invasive species (Mitchell & Power 2003; Torchin et al. 2003) (Fig. XIX.5).Only after a longer period of time are some parasites that occur in the given territory able to adapt to a new host sufficiently to survive consistently in the population of new hosts and are thus able to bring the spreading of the invasive species under control.
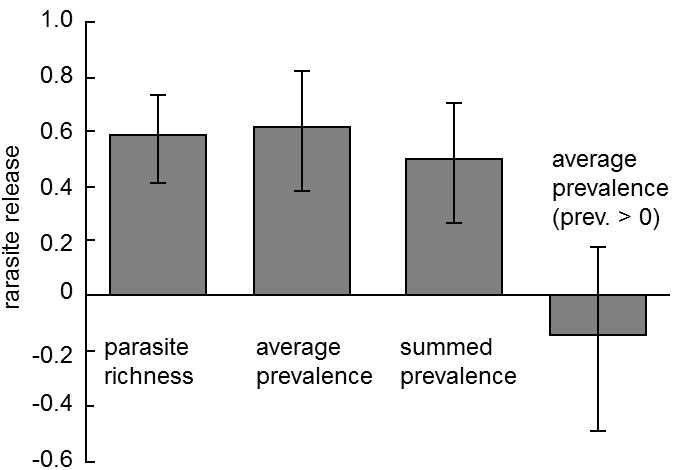
Fig. XIX.5 Escaping of introduced species from the reach of parasites. Species of animals that were successfully introduced outside of their original range managed to rid themselves from their parasitic burden to a great degree. On the ordinate of the graph are plotted the coefficients of freedom from parasites, calculated for the individual types of burden (parasite variety – number of species of parasite on the host species, prevalence – percentage of individuals in the host population uninfected by the given species of parasite, overall prevalence of the infection - sum of the prevalences for all the species of parasite) as (N – I)/N, where N is the parasite burden in the original range and i is the burden in the new range. It is apparent that the average number of species of parasite per introduced species decreased by approximately one half, and there was also a substantial decrease in the average prevalence of the individual infections (from 15% to 4%) and the average prevalence of all the infections together (from 133% to 71%). However, if only the species of parasite with non-zero prevalence were included in the calculation, i.e. species successful moved with the host to the new territory, there was no significant difference between the situations in the original and new ranges and, in fact, the average prevalence of the parasitic species was, to the contrary, higher in the new territory (increased from 23% to 28%). However, this higher prevalence only indicates that particularly those species of parasite that had a high prevalence in the original territory managed to get to the new territory. According to Torchin et al. (2003).
However, in some cases, the pathogenic manifestations of infection by an unadapted parasite are very drastic, so that in a great many cases it ends with the death of the host organism.These cases are apparently far rarer than cases where an unadapted parasite is not capable of reproducing at all and of substantially damaging the new host.However, from the viewpoint of humans, these cases are more obvious and thus they get far more publicity.If the population of hosts survives the meeting with the new parasite, changes gradually occur in the progress of the infection and, understandably, also in the dynamics of its spreading in the host population.Specifically, the pathogenic manifestations of parasitosis are reduced, so that the originally fatal infection is gradually reduced to a milder sickness and, in extreme cases, can even end up practically free of symptoms (asymptomatic) (Combes 1997).The well-documented history of the intentional introduction of the virus of rabbit myxomatosis into Australia (Fig. XIX.6) is a textbook case of reduction of the pathogenicity of infection that has been well documented.
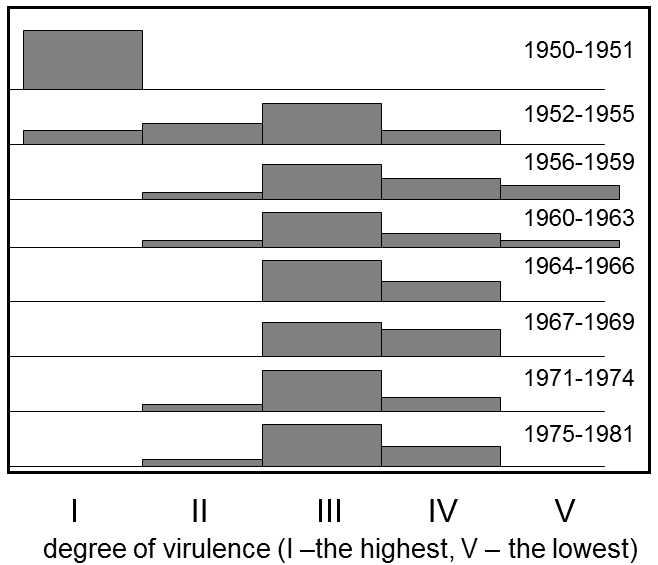
Fig. XIX.6 Changes in the virulence of myxomatosis viruses after introduction into Australia. It is apparent from the histograms that the greatest virulence (measured as the death rate of infected rabbits) was exhibited by myxomatosis immediately following introduction, i.e. in 1950, while the lowest was in 1956 – 1959. The virulence of the infection increased again at later dates. The dying out of the most virulent strains in the first years following introduction into Australia was probably a result of the fact that the viruses killed their hosts so fast that insect vectors were not capable of transferring the infection to another host. The dying out of the less virulent strains in the following years could have been caused by the fact that the prevalence of infection increased as the virulence decreased, leading to increased frequency of superinfections and, during them, the slowly multiplying viruses succumbed to competition from rapidly multiplying strains. According to Fenner (1994).
The phenomenon of gradual reduction in the pathogenicity of parasites, described commonly but rather inaccurately as the phenomenon of reduced virulence (see XIX.4.1), is, on the one hand, influenced by evolution of the host organism and selection of individuals that are more resistant to reproduction of the pathogen and to the pathological manifestations of its action in the organism and, on the other hand, selection within the parasitic species is important here, as parasites that do not excessively damage their hosts can produce more infectious stages and infect more hosts during their existence in a single host organism.
However, it should be pointed out that, in a great many cases, the pathogenic processes that accompany parasitosis are directly connected with the reproduction of the parasite – the more invasive stages it forms during the overall time of infection, the more intense are the pathogenic processes and their consequences (Fig. XIX.7).In this case, to the contrary, the pathogenicity gradually increases with gradually increasing adaptation of the parasite.Even cases where the pathogenic processes are part of manipulative behavior of the parasite and participate substantially in the effectiveness of spreading of the parasite in the host population are not exceptional (see below).Also in these cases, the virulence of the parasite is not reduced; to the contrary, the parasite is able to do more damage to the members of species or populations to which it is adapted in the long term (Ebert 1994; Ebert & Herre 1996)
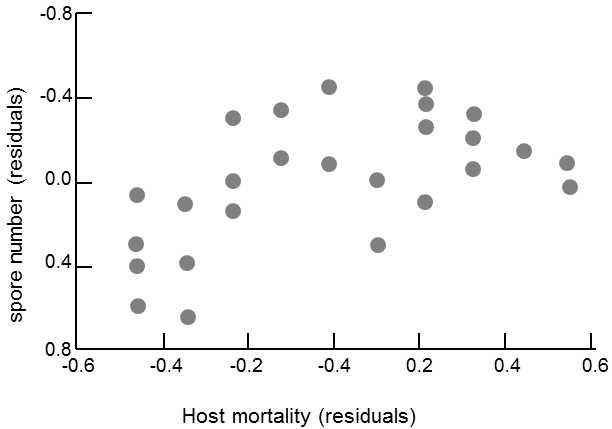
Fig. XIX.7 Correlation between the rate of reproduction of a parasite and mortality of the host. The number of spores of Pleistophora intestinalis (Microsporidae) formed on an average by the relevant strain of parasite in the digestive system of infected Daphnia magna is correlated with the mortality of the host. However, the dependence is not monotonous; this is a sharply positive dependence for strains forming a subaverage number of spores. However, the dependence tends to be negative for parasite strains forming a large number of spores in the host. The residuals were calculated by statistically eliminating the differences in sensitivity to infection in various host strains. Data from Eberta (1994), according to Eberta and Herrea (1996).
.
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.