XXV.4.2 When defining a particular taxon on the basis of shared traits, evolutionary systematists must take into account the definitions of other taxa within the relevant evolutionary line
An obvious requirement in defining taxa within a certain phylogenetic line is that taxa at the same level must not overlap and that certain species cannot be assigned to two various taxa at the same level. The only exception consists in species or phylogenetic lines that were formed, not by splitting of lines, but rather by fusion of species belonging to two different taxa. In this case, the relevant species and its descendants can be included in both the taxa from which the relevant parent species were derived and thus the taxon in which it finally ends up is only a matter of convention. Of course, it is more practical and more common to separate the descendants of such a fusion into a new taxon.
The entire system of overlapping taxa defined in the given phylogenetic line on the basis of the achieved level of anagenesis must also be created so as to reflect a certain hypothesis on classification of traits as homologies and homoplasies. In this respect, the system should be internally consistent. It is inadmissible to define a taxon so that its existence simultaneously necessarily requires the assumption that another taxon within the studied line is defined on the basis of homoplasy and is thus polyphyletic (Fig. XXV.7).
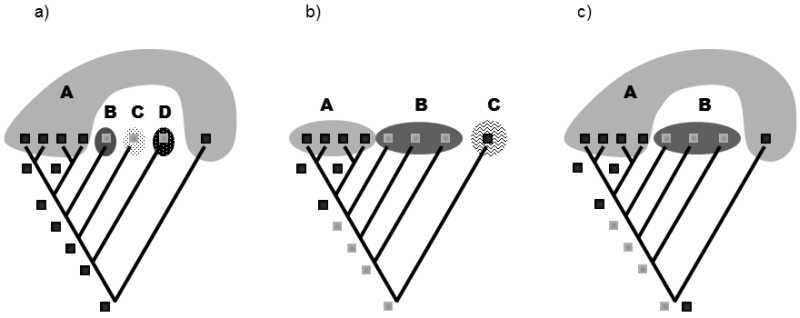
Fig. XXV.7 Creation of an internally consistent taxonomic system. Simplified phylograms depict cladogenesis and the distribution of the binary color trait (dark or light) with designation of three different variants of the taxonomic system. Variants (a) and (b) are internally consistent and thus permissible from the viewpoint of the principles of evolutionary systematics; variant (c) is not permissible, as it assumes that either taxon A or taxon B (depending on the form of the trait present in the common ancestor of the entire phylogenetic line) is defined on the basis of homoplasy.
Once again, it is necessary to bear in mind that all the taxa are, in actual fact, only our abstractions defined on the basis of subjective decisions. As individual taxa differ in their phenotypes, not in one, but mostly in a number of traits and, simultaneously, anageneses of the individual traits very frequently occurs independently and at various speeds, it need not necessarily be possible to create an internally consistent system of taxa based on the distribution of specific traits for existing species in a particular phylogenetic line. However, even this does not reduce the general requirements that the created system be unequivocal and not contain polyphyletic taxa. Our hypothesis about the classification of traits as homoplasies and homologies, on which the particular system is based, must, however, not contain any a priori inner contradictions.
Cladists generally hold the opinion that there is no difference between a polyphyletic and paraphyletic taxon that would be apparent directly from the topology of the cladogenesis scheme. This opinion has a certain realistic basis, as it holds that a paraphyletic taxon is a taxon defined on the basis of shared plesiomorphy, while a polyphyletic taxon is a taxon defined on the basis of shared homoplasy, where cladogenesis schemes do not, by definition, contain the relevant information related to anagenesis (otherwise they would be phylograms). In fact, however, it is possible to recognize the presence of polyphyletic taxa defined within a certain system even without knowledge of anagenesis and the distribution of the individual forms of the traits. In the absence of information on the distribution of the individual forms of the traits in modern species and their evolutionary ancestors, it is not possible to determine, on the basis of knowledge of cladogenesis, which of the defined taxa is paraphyletic and which polyphyletic; however, it is possible to determine that at least some of the defined taxa are polyphyletic and thus that the system as a whole is created incorrectly from the viewpoint of evolutionary systematics (see Fig. XXV.7c).
Draft translation from: Evoluční biologie, 2. vydání (Evolutionary biology, 2nd edition), J. Flegr, Academia Prague 2009.
The translation was not done by biologist, therefore any suggestion concerning proper scientific terminology and language usage are highly welcomed. You can send your comments to flegr cesnet [dot] cz. Thank you.
cesnet [dot] cz. Thank you.